
石油は、世界中で主要なエネルギー源を構成しています。世界人口の継続的な増加によるこの資源の過度の消費は、石油埋蔵量の枯渇につながりました。これは、温室効果ガスの放出による気候変動の影響の増大と一致しています (Stephanopoulos, 2007)。
これらの問題に対する潜在的な解決策は、トウモロコシ、ポプラ、スイッチグラスなどの植物から生成できる持続可能なエネルギーの代替源であるバイオ燃料にあります。
エンジンで使用される最初の主要な代替バイオ燃料は、1990 年代初頭に一般にエタノールとして知られていたバイオエタノールで、10% のエタノールと 90% のガソリン E10 燃料を作成するためにガソリンと混合されました。他のブレンドには、最大 15% のエタノールを含む E15 や、フレキシブル燃料車で使用できる E85 などがあります。今日、米国で使用されている燃料の 98% 以上が E10 混合燃料であり、これは一般的で便利な代替燃料となっています。
ただし、これは利用可能な最も効率的な再生可能燃料ではありません。エタノールは、バイオ燃料の使用に先行する確固たる出発点を提供してきましたが、この特定のバイオ燃料には、貯蔵エネルギー量が少ないことや、エンジン部品の製造に使用される鋼に亀裂をもたらす腐食効果など、燃料経済へのさらなる拡大を制限する固有の課題があります。 (クレメンテ、2015)。
その結果、研究者は、一般的なバクテリア Escherichia coli で遺伝子工学を使用して、より効率的な代替バイオ燃料を最適化しようとしました。 .私たちのほとんどにとって、E.大腸菌 食品汚染のニュース報道でのみ言及されています。遺伝子工学の世界では、大腸菌は、よく知られている成長代謝と利用可能な遺伝子工学ツールの最大のセットを備えた貴重なユーザーフレンドリーな宿主です。多くの人は、この細菌が広範囲に研究されていることを知って驚くかもしれません 遺伝子の調節と発現、および糖尿病治療のためのインスリン産生など、人間が使用するための貴重な物質の供給源として役立ってきました。 大腸菌 さまざまな糖を消費することができ、実験室規模と工業規模の両方で多様な成長条件を可能にし、E.大腸菌 遺伝子工学に最適な生物。
遺伝子工学の進歩により、遺伝学者は遺伝子を改変して、イソプロパノールなどの特定の潜在的なバイオ燃料を多かれ少なかれ生産できるようになりました。これらの変更は、新しい遺伝子を導入するか、既存の遺伝子を変更することにより、生物の遺伝子構成を変更します。これは、DNA 塩基対 (アデニン-チミン/グアニン-シトシン) の変更、遺伝子全体の削除、または複数の遺伝子の挿入によって行うことができます。生物工学者が代謝経路全体をある生物から別の生物に移すことを可能にするツールが開発され、科学者が実験室でより簡単に操作された生物を扱うことができるようになりました。これらのツールは一般に、ウイルスやバクテリアなどの生物で使用できます。遺伝子または遺伝子の配列を別の生物に導入すると、この生物はイソプロパノールなどの目的の分子を生成できます。
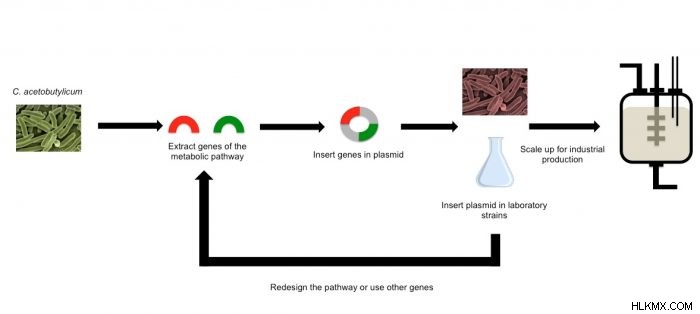
アセトンとイソプロパノールは持続可能で、安価で、エネルギー密度が高く、代替燃料としてテストされています。これらのバイオ燃料はすでに既存のエンジンと互換性があり (Poh &Poh, 2017)、イソプロパノールはエタノールよりも混合能力が高く、腐食効果が少ない.イソプロパノールと同様に、アセトンはエタノールに比べてエネルギー密度と混合能力が高く、エンジンに対する腐食性も低くなります。これらの特性はすべて、イソプロパノールとアセトンの両方がバイオ燃料市場でエタノールを補完または代替する可能性を秘めていることを意味し、これらは両方とも Clostridium acetobutylicum と呼ばれる細菌によって自然に生成されます。
クロストリジウム・アセトブチリカム は、単一の代謝経路を介してイソプロパノールとアセトンを生成します。つまり、この生物は、同じ遺伝子配列を使用してこれらの潜在的なバイオ燃料の両方を生成します。この遺伝子経路の一次生成物としてアセトンが生成され、副生成物としてアセトンからイソプロパノールが得られます。この経路は、これら 2 つの分子の生産を最大化するようには設計されていませんが、そうするように設計することはできます。残念ながら、C で使用できる遺伝子工学ツールは.アセトブチリカム は限られており、バイオ燃料の収量を最適化するには、他のいくつかの生化学的経路を阻害する必要があります (Toogood &Scrutton, 2018)。
幸いなことに、Cのアセトン経路。アセトブチリカム Eに転送できます。大腸菌 Cの制限なしにアセトンおよびイソプロパノールの生成をもたらす。アセトブチリカム。 遺伝子工学者は、E でアセトンとイソプロパノールを生成するために使用される遺伝子を「過剰発現」させることができます。大腸菌 これは、これらの遺伝子をオーバードライブ状態に押し込んで、潜在的なバイオ燃料用のイソプロパノールとアセトンを大量に生成できることを意味します (Atsumi &Liao, 2008; Koppolu &Vasigala, 2016)。
遺伝子工学の進歩により、科学者は生物の自然な代謝経路を研究および改善して、人間が健康、エネルギー、食物の利益のために大量生産できる化合物を生成できるようになりました。 C.アセトブチリカムから代謝経路全体を移す能力 大腸菌への変換は、従来の燃料を改善または置き換えることができる効率的で高品質のバイオ燃料を生産する生物の自然な能力の商業的に実行可能なアプリケーションにつながる可能性があり、それによってエタノールに関連する新たな供給と環境問題を緩和します。排出量を削減し、エネルギー収量を改善するためのエンジンと燃料混合物の最適化には、まだ課題が残っていますが、現在の結果は、石油への依存を減らすという私たちの使命を後押ししています。
これらの調査結果は、テネシー大学ノックスビル校の Bredesen Center for Interdisciplinary Studies の Jessica Velez、Laval University の Institute of Integrative and Systems Biology の Angel Fernando Cisneros Caballero、および Division of Immunity の Narjes Alfraiji によって行われた研究に基づいています。マンチェスター大学の感染症および呼吸器内科。
参考文献:
<オール>






