IPBES (1、セクション 3.2、pp. 196 を参照) が最近再び述べたように、生物多様性の保全は人類にとって重要な課題です。最適な生物多様性の原則 (2, 3) は、生物学的基礎に戻り、なぜ自然が多様性を必要とするのかを理解しようとする試みです。
多様性が適応である場合
生物学的システムの内部の多様性 (つまり、構成要素の多様性) は、その有効性、回復力、および実行可能性に関連する適応特性であると考えています。最適な多様性を備えた生物学的システムは、最も効果的で実行可能です。最適な多様性から離れると、生物学的システムの有効性と実行可能性が低下し、最終的にはそれを破壊することさえあります.
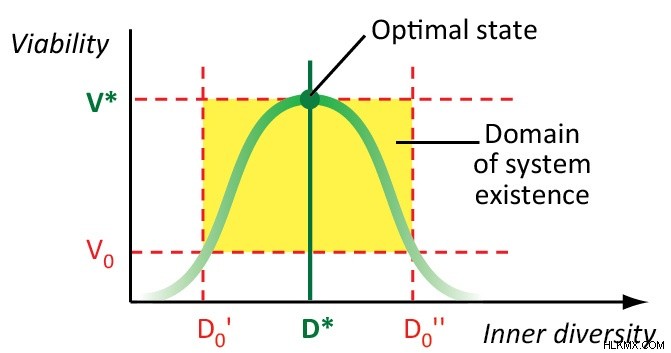
最適な生物多様性は、さまざまな階層レベルの生命の相互作用の間に確立されます。私たちの理論モデル (2, 3, 4, 6) は、生命階層の小さな断片、つまり、個体群とそれらで構成されるコミュニティ、つまり個体群内と種の多様性のみを研究しました。これらのモデルからさらに考えが生まれます。
人口とコミュニティ レベルの反対の反応 – 同じコインの表と裏
個体群内と種の多様性の最適値は、環境パラメーター (利用可能なリソースの量と安定度) に逆モードで依存します。最適な個体群内多様性は、より安定した条件では減少しますが、最適な種の豊富さは増加します。最適な種の豊富さも、より「豊かな」条件で増加し、最適な個体群内の多様性は資源の量に依存しません。個体群内と種の多様性は、生物学的階層の 2 つの不可分な側面を示しており、それらの反応の違いは、適応プロセスにおける特定の役割を示しています。
ボトムアップの最適化:コンポーネントは上位レベルで多様性を定義します
最適な多様性の値は、次の個体群パラメータに依存します:
– 個体の生態学的許容範囲の幅;
– 最大個体数増加率と死亡率;
– 基本資源コスト表現型と許容範囲拡大のコスト.
個体群の生存率を高めるこれらの各パラメーターの改善は、環境の安定化と同じように、最適な多様性の変化につながります。したがって、個体群は環境の不安定性を補うさまざまな方法を持っています。個体群の増加率の増加、死亡率の低下、個々の許容ゾーンの拡大、表現型のリソースコストの低下などです。これらのメカニズムは、その適応能力により個体群内で機能し、種構成の変化によりコミュニティ内で機能することができます。たとえば、K-ストラテジストとr-ストラテジストの間、およびスペシャリストとジェネラリストの間でシフトします.
上記のパラメーターのいずれかが改善され、他のパラメーターは変更されない場合は、生物の進化レベルが向上したと解釈できます。したがって、最適な個体群内の多様性が減少し、最適な種の多様性が増加する可能性があります。
最適な生物多様性の予想パターン
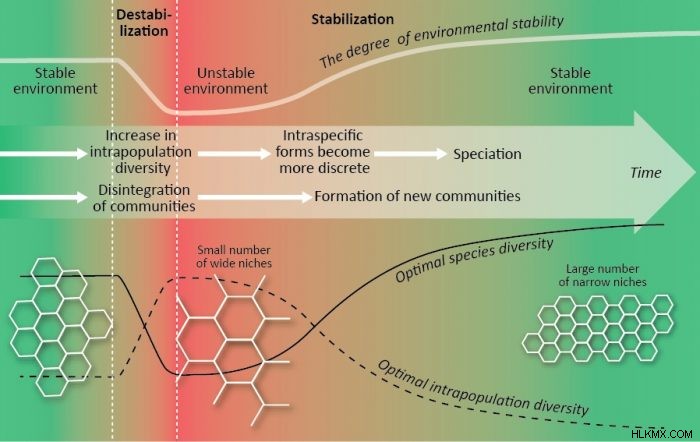
個体群内の多様性は、他の条件が等しい場合、生態学的ニッチの幅の代理と見なすことができます。したがって、群集の最適な生物多様性構造を予測することができます:1) 豊かで安定した条件で狭い生態学的ニッチを持つ多数の専門種、2) 乏しい不安定な条件で広い生態学的ニッチを持つ少数のジェネラリスト種、3)中間条件における中間最適ダイバーシティ構造 (6)。邪魔されていないクライマックス コミュニティとその構成集団は、最適な多様性に最も近いようです。いくつかの一致する生物多様性パターンは、地球規模と景観規模の両方で見つけることができます (7)。
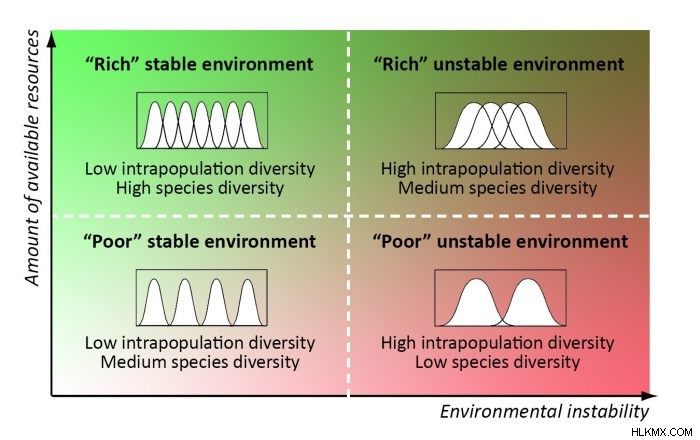
自然管理の実践では、これは、保全の優先順位の基準は、最適な多様性から人為的変化の距離であるべきであることを意味しますが、不安定または希少な条件に適応したコミュニティは種の多様性が比較的低いため、形式的な多様性指数(種の豊富さなど)は高くありません。それにもかかわらず、最大限の機能を保証します。
最適性の永遠の追求
生物系は最適な多様性を達成できるか?私たちの変わりやすい宇宙ではありそうにありません。環境または生物系自体の自然または人為的な変化は、それらを最適な状態から次善の状態にシフトします。したがって、生物系は、生態学的、小進化的、および進化的プロセス (同時に発生する可能性があります) の間、最適な状態を永続的に「追求」しています。
生態学的な例 1:共生生物相における r 戦略家の優位性
環境の人為的変化の一般的な方向性は不安定化であり(7)、集団に対する人間の影響は集団内の多様性の減少で表されます。後者は、集団が不安定化に適応する可能性を減らします。その結果、適応が群集レベルで機能し始め、典型的な在来種が他の種に置き換えられ、種の構成が K 戦略家から R 戦略家へ、専門家から一般主義者へと変化します。これは共生生物相の増殖に対応します。
生態系の例 2:コミュニティ アセンブリのメカニズム。
コミュニティの組み立てにおける特定のメカニズムの普及は、環境の「豊かさ」と安定性の比率、または特定の量のリソースと特定の範囲のリソースの下で存在できる最適なニッチの数の比率によって決定されます。環境パラメーター (例:t °C) の変動 (6):
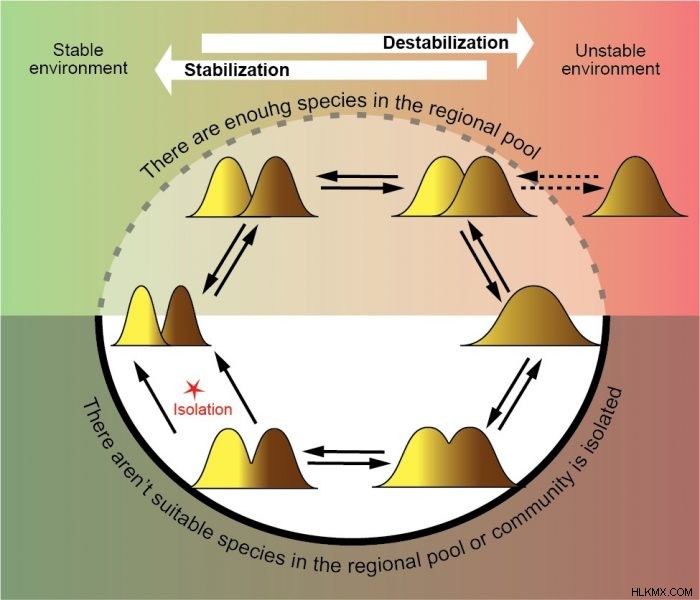
小進化の例:多様性最適化のメカニズムとしての同所的種内形態。
環境の安定度が変化すると、群集はさまざまな最適な状態の間を移動する必要があります。つまり、ニッチが狭い安定した状態の多数の種と、ニッチが広い不安定な状態の少数の種です。地域プールに十分な数の種があり、コミュニティが最適性を達成するために必要な特性を持つ種を簡単に含めることができる場合、問題はありません。しかし、コミュニティが孤立している場合や、地域のプールに適切な種が存在しない場合はどうなるでしょうか?この場合、1 つの種が複数のニッチを占有する場合、同所性種内形態 (エコタイプ) の形成により最適化が発生する可能性があります。このプロセスは、種内多様性 (ニッチ幅) と群集多様性 (種数) の両方を最適化できます。同所性種内形態は動的システムとして機能し、形態の分離が新しい種の形成につながるまで、地域の生物多様性を環境の変化に絶えず適応させます (3)。
進化の例:浮き沈みはあるものの全体的に多様性が増しています。
生物多様性の最適化は、V. Zherihin の生物セノティックな進化の調節の概念と V. Krasilov のコヒーレント - インコヒーレント進化の概念のプロセスの追加の説明である可能性があります。環境の不安定化とコミュニティの破壊 (生セノティック危機) の歴史的な期間は、個体群内の多様性の増加、ニッチの拡大、特殊な種の絶滅、種の多様性の低下によって特徴付けられます。個体群内の多様性の増加は、種内形態および種分化の将来の形成のための材料を提供します。環境の安定化と「新世代」のコミュニティの開発の歴史的な期間は、個体群内の多様性の減少、ニッチの狭小化、種内形態の離散化、種分化、種数の増加によって特徴付けられます。生物の進化レベルの増加は、個体群内の多様性の減少 (最適なニッチが狭くなった) と最適な種の多様性の増加 (上記参照) につながります。生命の進化 (3).
参考文献:
1) 最適な「リソース」ニッチの数と最適な「範囲」ニッチの数がほぼ等しい場合 (R/R *≈D/D*);
2) 中立的なメカニズムは、非常に「豊かな」環境で主に機能します。この環境では、大量のリソースにより、環境パラメーターの範囲よりもはるかに多くの種が存在できます (R/ R*>> D/D*);
3) 「非生物的フィルター」は、主に不毛の (過酷な) 環境で機能します。この環境では、少量の資源で、環境の範囲よりもはるかに少ない数の種の存在が可能になります。パラメータは (R/R*<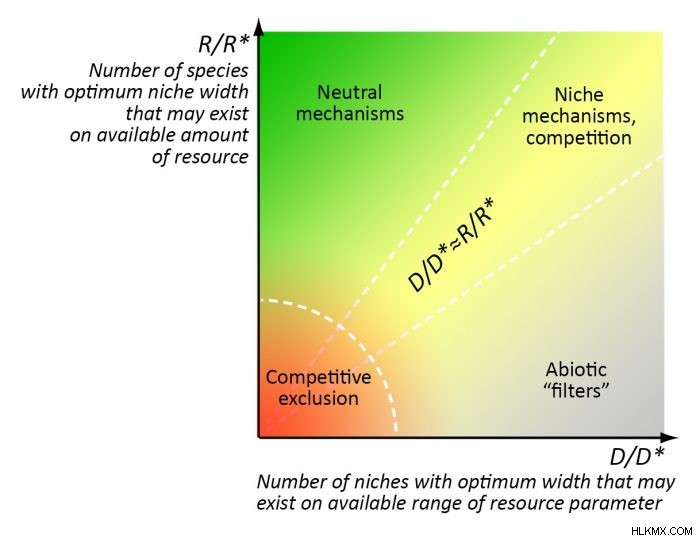
https://ipbes.net/sites/default/files/2018_eca_full_report_book_v5_pages_0.pdf
https://sciencetrends.com/
https://link.springer.com/article/10.1134/S1062359010010012
http://article.sapub.org/10.5923.j.ije.20120204.06.html
http://www.sciencepublishinggroup.com/journal/paperinfo.aspx?journalid=118&doi=10.11648/j.ajls.20130104.16
https://www.sciencedirect.com/science/article/abs/pii/S1470160X17302170?via%3Dihub