顕花植物が白亜紀前期 (1 億 3000 万から 9000 万年前) に地球上に出現して以来 (1)、それらは非常に複雑な生物地理学的プロセスを経て進化し、多様化してきました (2)。それらのいくつかは一連の系統に特有のものでしたが、他のものは主に広範に広がっていました.
しかし、研究者は現在、博物館や植物標本館で大量に文書化された知識に基づいた実際のシナリオしか見ることができません。これまでのところ、さまざまな植物種のセットが「空間的に構造化」されており、種の分布の優れたモザイクを形成しています。その後、生物地理学は、主に 19 世紀に、このような分布のパズルに取り組む生物科学の分野として登場しました。したがって、生物地理学者は、比較および分類の観点から、過去、現在、および未来の空間パターンを記述して、それらのパターンを持っている、持っている、そしてそれらのパターンを生成するプロセスを完全に理解することを試みてきました.
植物の種だけを考えると (ただし、動物の話は非常に似ています)、6 つの主要な生物地理学的領域があり、それぞれに固有の植物群構成のセットがあることがわかっています (3)。南アメリカ南部とメキシコ北部の一部を除いて、その境界がラテンアメリカとほぼ正確に一致する新熱帯地域には、地球上で大多数の種があります。少なくとも 40,000 種の顕花植物が固有のもの、つまり固有のものです。エリアへ (4)。ブラジルは、西側世界最大の国の 1 つであり、ラテンアメリカ最大の国の 1 つであり、この膨大な多様性の主要部分を占めています。この国には、33,000 種以上の顕花植物が、さまざまな森林またはサバンナのような生物群系で発生しています (5 ).
新熱帯区のジオバイオティック シナリオも複雑です。過去数百万年の間に多くのことが起こりました。アンデスの隆起、パナマ地峡の隆起、流量の点で世界最大の排水システムであるアマゾン川 (6) への変化。そして、アマゾンと大西洋の森林によって形成された古代の南アメリカの熱帯雨林の回廊が、開放的で季節的に乾燥した植生に置き換わった (7)。このような地質生物学的出来事は明らかに、地域の動植物に影響を与えており、時には異なる方法で、時には出来事が独自の共有された歴史を明らかにするような方法で.
私たちの調査の焦点である熱帯雨林の場合、いくつかの特定の出来事が、これまでに記録した生物相の分布に貢献したと考えています.それでも、これは、一度特定され、さらに説明されたパターンのためにのみ可能です.したがって、世界の特定の生物学的地域のアイデンティティを設定するには、一貫した分類システムでそれらの新しい地域を記述して適切に分類すること、および多くの固有種がある地域を推定できる新しい方法を記述することが含まれます。実際のところ、これらの生物領域を「固有の領域」と呼ぶことができます (8)。
とはいえ、研究モデルとしてミカン科の顕花植物 (オレンジ、グレープフルーツ、ライム科) を使用して、生物地理学的パターン認識の調査を行いました。このグループは新熱帯地方で特に多様であり、多くの種は私たちがパターン研究に焦点を当てた森林地帯に集中しています。そこで、この家族の 1 つの部族、ガリペイナエを選択しました (図 1)。ガリペイナ亜科には 28 の属と c があります。 130 種が記載されており、新熱帯地方の固有種です。それらの代表者のほとんどは、大西洋岸森林の特定の部分だけでなく、アマゾンや、ブラジルのセラードなどの開放的な季節的に乾燥した植生にも固有のものです。したがって、このグループは、ガリペイナエ自体の自然史だけでなく、熱帯雨林の生物地理学と進化に新しい洞察をもたらすための優れたモデルとして役立つことができると私たちは主張します.

次に、ガリペイナエ種のすべての分布データを手動で大幅に改訂しました。入手可能な文献の大規模な評価と、植物標本館や博物館に寄託された標本の再検討を通じて、各発生記録の地理的座標を特定、修正、または選択し、それらの分布をマッピングしました。次に、それぞれの分布図を作成し、ブラジルで2つ以上の固有種の分布パターンが似ている場所を調べました。これらは後に、調査地域のグループのいわゆる「固有地域」として定義されました。
要約すると、大西洋岸森林におけるガリペイナエの固有種の発生の 3 つの「ホット スポット」が見つかりました (図 2)。言い換えれば、Galipeinae 種はブラジル東海岸のさまざまな緯度に無作為に分布しているわけではなく、代わりにこれら 3 つの部分に限定されたいくつかの固有種が存在します。興味深いことに、これらの特定された固有の領域は、植物と動物 (収穫者、鳥、有鱗目) の両方に関する他の研究の他のグループについても記録されていました。問題は、大西洋岸森林のこれら 3 つの地点での分布の収束をどのように説明できるかということです.
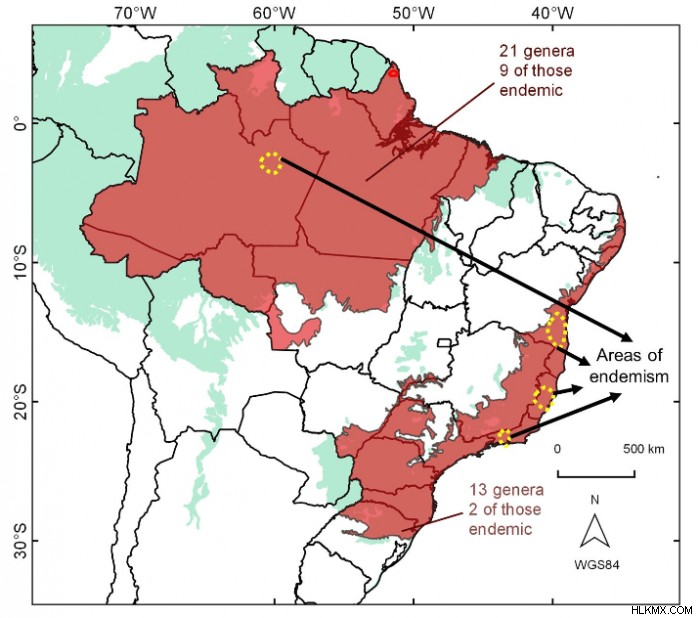
これは実際には難しい質問ですが、私たちの論文では基本的に 2 つの古典的な仮説について説明します。 1 つ目は、森林避難仮説 (6, 9) です。それは、更新世後期 (126 から 11.6 千年前) 以来、気候変動が継続的な森林の収縮と後退を引き起こしたと述べています。言い換えれば、より乾燥した寒い時期には、湿度に依存する森林生物の分布が狭くなり、代わりにサバンナのような植生が拡大する可能性があります.
逆に、湿度が高く温暖な時期には、最も湿潤な地域が再び拡大し、かつて破壊された常緑樹林の孤立した「避難所」が再接続されます。したがって、このような「常緑樹林の避難所」は、地元の動植物を分断する可能性のある場所になります。したがって、いくつかの継続的な個体群はしばらくの間孤立し、いくつかの突然変異を蓄積して進化し、最終的にそれらが再接続されたときに、2 つの新しい別々の系統を区別できるようになります:1 つの姉妹がもう 1 つの姉妹であり、それぞれが森の特定の場所に固有のものです。 .私たちはそのような自然現象を「ヴィリアント種分化」と名付け、それぞれの避難所は、かつては連続していた系統の種分化プロセスにおいて「ヴィリアント エージェント」としての役割を果たします。
確かに非常に便利な仮説ですが、その反論の証拠をいくつかまとめました.特定のグループに関するいくつかの新しい生物学的データと、南アメリカに関する新しい地質学的証拠は、ほとんどの森林系統の年齢が、そのような変動が起こった後期更新世よりもはるかに古いことを示しています(10)。そのため、別の仮説が浮上しており、種分化の新しいバイカリアント エージェントが示唆されています。大きくて幅の広い川はその 1 つであり、アマゾン川の排水システムの歴史的な変化を考えると、特にアマゾンの血統にとって重要な役割を果たしている可能性があります (6)。大西洋岸森林については、別の、さらに示唆に富んだ仮説が提案されています。「アトランティス森林仮説」です (11)。この仮説は、大西洋における定期的な海の後退と海進が、大西洋岸森林のさまざまな部分に氾濫し、系統を海に囲まれた「島」に分割したと述べています。しかし、この考えは批判も受けています (12)。
結局、ガリペイナエに何が起こったのか、まだはっきりとは言えません。実際のところ、代替シナリオの 1 つは、これらすべての仮説が排他的ではないため、一緒に作用した可能性があるというものです。しかし、分布データと文献からすでにわかっていることだけで、私たちがすでにできることを調べて仮説を立ててください。たとえば、新しい生物学的 (系統発生) および地質学的 (気候、エダフィック) データを組み込んだダウンストリーム分析に直接進むことができるようになりました。ガリペイナ科のこれら 3 つの主要な固有性領域が回復したので、次のステップは、南アメリカの地球生物史とさまざまなグループのより完全なパノラマに向けて、各候補仮説をテストすることです (または、新しいシナリオを提案することさえありますか?)。開花植物。
参考文献:
<オール>