ほとんどの複雑な生物は、人生のある時点で奇妙なゲノム計算を行います。増殖するために、減算してから加算します。つまり、減数分裂のプロセスを通じて生殖するために、通常の半分の数の染色体を持つ特殊な性細胞または配偶子を作成します。次に、それらの配偶子のペアをマージして、完全で一意のゲノムを持つ新しい個体を作成します。有性生殖は真核生物の間でほぼ遍在しています — ケルプからコアラまで、膜で結合した核とオルガネラを持つ生物です。私たち動物は精子と卵子を融合させます。きのこは、菌糸の地下衝突から発芽します。花粉は花の組織を駆け抜ける管を送って胚珠に結合し、それらが出会う場所で果実と種子を作ります。
しかし、このように細胞を融合させることは、通常の細胞のライフ サイクルに反します。細胞は有糸分裂によって 2 つに分裂して無性生殖を行いますが、それ以外の場合は、ほとんどの場合、系統を消滅させる可能性のある完全性の大きな混乱を防ぎます。
この危険な取り決めが進化の過程で取り込まれた理由は、熱心な研究の対象となっています。一般に、有性組換えは、種のゲノムを健康に保ち、リスクを相殺するのに十分なほど多様であると考えられています。しかし、研究者たちは、それがどのように進化したかについての分子の手がかりもつなぎ合わせています.
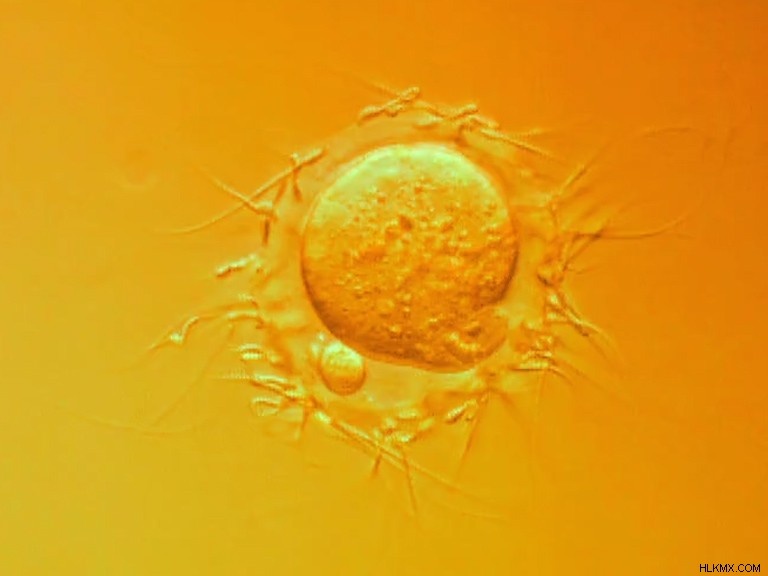
最近、biorxiv.orgプレプリントサーバーで共有された研究は、一部の細胞が進化の歴史のはるか昔に膜を融合させる力を持っていたことを示唆しています.有性生殖のこの部分を可能にする分子機構は、20 億年以上前に古細菌と呼ばれる単純な原核細胞に存在していた可能性があり、おそらく真核生物と性が進化する 10 億年も前に存在していた可能性があります。しかし、新しい発見は、有性生殖のためのこの種の細胞融合が、生命の歴史の初期に現れなかった理由の説明も示唆しています.
幅広い融合タンパク質ファミリー
真核生物の系統樹全体で、同じタンパク質が配偶子の細胞膜と核膜の融合を可能にします:HAP2.それは真核生物の間で非常に普遍的に共有されているため、「共通の祖先は同じメカニズムを持っていたに違いない」とイスラエルのヘブライ大学エルサレムの細胞生物学者であるマイケル・ブランダイスは述べた.
しかし、この最後の真核生物の共通の祖先は、タンパク質の最初の担い手ではなかったかもしれません. HAP2 は、一部のウイルスがウイルスエンベロープを細胞膜と融合させるために使用するタンパク質のクラスを含む「フセキシン」のスーパーファミリーに属しています。進化生物学者にとって、HAP2 とこれらのウイルス フセキシンとの関係は、「ウイルスか卵か」という謎を突きつけます。最初のフセキシンは古代の真核生物で進化し、ウイルスによって独自の目的のために盗まれただけなのですか?それとも、フセキシンはある時点で真核生物が配偶子融合のために採用したウイルスの発明だったのでしょうか?すべての真核生物およびウイルス誘導体が進化したタンパク質であるフセキシンの最も古い形態を見つけることは、これらの答えを学ぶために重要です.
古細菌は、答えを探す自然な場所でした。それらは真核生物の祖先であると考えられているため、ミトコンドリアと葉緑体になった共生細菌の最初の宿主であったため、古細菌は真核生物の系統樹の根底にあるものを示しています。さらに、研究者は、減数分裂中の遺伝物質の管理と組換えに関与する遺伝子の起源を古細菌にまで追跡しました。 (古細菌は、これらの遺伝子の特別な能力を使用して他の遺伝子を修復しているようです。) したがって、古細菌は、イスラエル、スウェーデン、アルゼンチン、ウルグアイ、および新しい研究の背後にある他の国の科学者の国際協力が答えを探し始めた場所です.
イスラエルのハイファにある Technion の Benjamin Podbilewicz、スウェーデンの Karolinska Institute の Luca Jovine、アルゼンチンの生理学、分子生物学、神経科学研究所の Pablo Aguilar、および彼らの同僚は、古細菌の仮説的フュキシンがどのように見えるかを予測しました。現代の真核生物におけるHAP2の構造とタンパク質配列。ゲノムデータベースのスキャンを含むさらなる研究により、予測されたフセキシン構造に驚くほど似た形状のタンパク質を生成する可能性が高いと思われる96の古細菌遺伝子が発見されました。研究者はこれらの古細菌遺伝子をfsxAと呼んだ .
チームは、古細菌のゲノムで古代のフセキシンを発見したように見えました。これは、生命のその領域で初めて見られたものです。しかし、このタンパク質が生きた細胞系でフセキシンのように振る舞うという保証はありませんでした.
Podbilewicz と彼の同僚は、これらの遺伝子の囁かれた可能性に命を吹き込みました。彼らは fsxA を組み込みました 培養ハムスター腎臓細胞に導入し、遺伝子の発現を可能にしました。一部の細胞を蛍光赤色の核を持つように変更し、他の細胞を蛍光緑色の核を持つように変更することで、研究チームは細胞が推定上の古細菌のフセキシンを使用して融合しているかどうかを確認できました。
そして、それはまさに彼らが見たものです。 fsxA 遺伝子は配偶子のペアであるかのように融合していました.
古細菌に機能的なフセキシンが存在することは、性のための分子ツールが真核生物の進化よりも前に存在していたことを意味している可能性がある、と研究者は示唆している。遺伝子の進化的分析に基づいて、彼らは、これらの古細菌のフセキシンは、これまでに見られたタンパク質の最も原始的な形態であり、数十億年前のものである可能性が高いと考えています.
融合前の機能
この研究には関与していない、ブラジルのサンパウロ大学の進化生物学者である Paulo Gonzalez Hofstatter は、この発見が進化の歴史に関する私たちの知識のギャップを埋めていると考えています。 「しかし、ある意味では、それは少し予測可能でもあります」と彼は言いました。
それはまた、フセキシンの起源に関する最終的な言葉ではないかもしれません. Hofstatter は、最古のフセキシンがウイルス性であった可能性は依然としてあると警告している。ウイルス性タンパク質の遺伝子は古細菌に移行し、その後真核生物に移行した可能性がある.
フセキシンが本当に古細菌の革新であったかどうかにかかわらず、新しい研究はこれらのタンパク質の本来の機能に関する重要な疑問を提起します.
ユニバーシティ カレッジ ロンドンの進化遺伝学者である Andrew Pomiankowski は、FsxA タンパク質が哺乳動物細胞で発現されると細胞融合を可能にするように見えるが、これらの意図されているフューキシンが古細菌で何をしているのかはまだ不明であると述べています。極度の熱や圧力などの厳しい環境条件を必要とすることが多いため、古細菌は実験室で培養するのがはるかに難しい傾向があるため、バクテリアよりも古細菌の内部の仕組みを調べるのは困難です.
それにもかかわらず、彼はポドビレヴィチと彼のチームがとったアプローチを称賛します。 「彼らはまさに正しい質問を提起しています。この[融合]遺伝子は原核生物で一体何をしているのですか?」彼は言った.
1つの可能性は、古細菌が性的プロセスに大まかに対応する一種の遺伝子交換を行うのを助けることです. Hofstatter は、古細菌のいくつかの種は、漠然と減数分裂に似た細胞融合と DNA 組換えの限られた形態を実行することを指摘しました。 「それらは融合し、再結合し、再び分裂するだけです」と彼は言いました。
横移動の代替品
フセキシンが古細菌から真核生物にどのように移行したかについても明らかではありません。ホフスタッターにとって最も可能性の高い説明は、すべての真核生物を支える古細菌と細菌の共生において宿主細胞となった系統への古細菌からの直接的(または「垂直的」)な遺伝である.本質的に、真核生物はこのように生まれた可能性があり、祖先からの古いトリックを転用しています.
しかし、この説明には注意が必要な理由があります。イリノイ大学アーバナ シャンペーン校の微生物学者 Rachel Whitaker は、研究チームが塩分を好む古細菌である Halobacteria のグループにフセキシンを発見したが、Crenarchaeota のグループには発見しなかったことに驚いている。 Halobacteria よりも真核生物です。
研究者によって発見された潜在的に重要な詳細の 1 つは、fsxA です。 遺伝子は、統合されたモバイル要素として知られる古細菌ゲノムの特別なセクション内に存在し、ゲノム内を動き回ることができ、時には種の壁を越えて他の個体のゲノムに移動することさえできます.
これらのフセキシン遺伝子が実際に少し機敏である場合、最も初期の真核生物が古細菌細胞からこれらのジャンピング融合遺伝子を取り込み、後に「exaptation」の例における配偶子融合における役割のために遺伝子を改造した可能性があります。古い特性の新しい用途。かつては古細菌細胞間の限られた遺伝的伝達に使用されていた可能性のあるフセキシンは、有性生殖の進化の種をまいた可能性があります。 (Podbilewicz と彼の同僚は、彼らの研究の査読が保留されているため、この記事のインタビューを拒否しました。)
いずれにせよ、古代の古細菌がおそらくこれらの融合タンパク質を持っていたという発見は、真核生物の進化の理解に影響を与えます.確かに、原核生物で性的細胞融合がうまくいかなかった理由についての手がかりを提供するかもしれません.解決策は、古細菌が他の遺伝子や環境から新しい遺伝子を収集する方法に見出される可能性がありますが、原始フセキシンが可動性の遺伝要素に存在するという事実にも見出される可能性があります.
Pomiankowski は、古細菌のような原核生物は、ウイルス感染やプラスミド (細胞間を移動できる遺伝物質の断片) を介して遺伝子を交換できると説明しています。しかし、彼らは環境から DNA を拾い上げ、それをゲノムに再結合することもできます。これは、形質転換と呼ばれるプロセスです。これらの遺伝子交換の慣行は、潜在的に有用な新しい遺伝子を導入し、有害な遺伝子変異体の蓄積を抑制することにより、古細菌のゲノムを維持するのに役立つ可能性がある、と Pomiankowski は述べた.
しかし、古細菌とバクテリアが真核生物として恒久的に力を合わせると、それらの細胞はますます複雑になり、それらのゲノムは急速にサイズが大きくなりました.その結果、古い「水平遺伝子導入」戦略は支持できなくなった可能性があります。
「横方向の遺伝子導入の欠点は、ゲノムが大きくなると、ますますうまくいかなくなることです」とポミアンコフスキーは言いました。 「[もし] 環境からほんの少しの DNA を選んだとしたら、それが一致する確率はどのくらいですか?それはゲノムサイズの減少関数です。」
再利用された先祖のフセキシンによって可能になった細胞融合は、より協調的で大規模な遺伝子交換、つまり有性生殖を可能にすることで、真核生物への移行に貢献した可能性があります。このような根本的なシフトは、生まれたばかりの真核生物のゲノムを維持するのにより適切だった可能性があります。
答えを持つ生物
別の可能性は、主に真核生物自体の利益のために配偶子融合が進化したのではないということです。むしろ、野心的な遺伝子の操作的な自己保存の目的に役立った可能性があります。古細菌のフセキシンは可動性の遺伝要素と非常に密接に関連しているため、ポミアンコフスキーは、フセキシンは「それ自体の利益のために細胞間の融合を引き起こしたいある種の要素」である可能性があると考えています。統合された可動要素の助けを借りて、フセキシン遺伝子は、最初に原核生物の融合を介して、次に真核生物の融合と遺伝子交換によって、それ自体の増殖を促進した可能性があります.
しかし、Pomiankowski は、この理論が古細菌に統合された可動要素が本物のジャンパー遺伝子であったことにかかっていることを認めており、これは不確実である.このような転位因子は、原核生物ではほとんど見られません。人間の場合、「あなたは 50% 転移可能な要素です」と Pomiankowski 氏は述べています。 「でも、バクテリアの場合は 0.5% くらいです。」
細胞融合は有性生殖の重要な部分ですが、古細菌における基本的な融合と精子と卵細胞の融合の間には、他にも多くの進化段階がありました。この進化的変化を調査することは、真核生物の最後の共有祖先にできるだけ近い生物を見つけて研究することを意味するかもしれません.
Hofstatter はおそらく、鞭毛虫と呼ばれる単細胞の真核生物をより詳しく調べることを意味すると考えています。鞭毛虫は非常に多様で水生生息地に豊富に存在し、真核生物の最後の共通祖先に非常に似ていると考えられています。 「性別の起源を理解する鍵は、これまで文化にないいくつかの鞭毛虫のサンプリングです」と彼は言いました.
注目すべきもう 1 つの場所は、ほんの数年前に記述された、真核生物の最も近い親戚であると考えられている、謎めいた古細菌のアスガルド系統です。彼らは、細胞融合やその他のプロセスがどのようにしてセックスを生み出し、活気に満ちた賑やかな真核生物のもつれをもたらしたのかという問題に、ある程度の解決策を提供できるかもしれません.