主な違い - 原核翻訳と真核翻訳
原核生物および真核生物の翻訳は、mRNA によって運ばれる遺伝的指示を解読することによって、タンパク質の合成に関与しています。翻訳中、mRNA 上のコドンとして知られるヌクレオチド トリプレットは、一連のアミノ酸に翻訳されます。原核生物と真核生物の両方の翻訳は、プロセス全体で同様の基本計画を共有しています。ただし、これらの翻訳プロセスにはいくつかの違いが見られます。 主な違い 原核生物と真核生物の翻訳の違いは、原核生物の翻訳はその転写と同期して起こるのに対し、真核生物の翻訳はその転写と非同期に起こるということです。
この記事では、
1.原核翻訳とは
– 定義、プロセス、機能
2.原核翻訳とは
– 定義、プロセス、機能
3.原核生物と真核生物の翻訳の違いは何ですか
原核翻訳とは
原核生物では、翻訳は転写と同時にタンパク質を合成するプロセスです。翻訳は、遺伝子の 5' 末端が mRNA に転写された直後に始まります。原核生物の翻訳は基本的に、開始、伸長、終結の 3 つの段階で行われます。翻訳を開始するために、2つのサブユニット50Sおよび30Sが組み立てられる。 3 つの開始因子、IF1、IF2、および IF3 は、開始複合体を組み立てるのに役立ちます。 N-ホルミルメチオニンは、翻訳で最初に付加されるアミノ酸です。 GTP は、残りのヌクレオチドと入ってくるヌクレオチドの間のペプチド結合形成のエネルギー源として使用されます。翻訳開始因子はEF-Pです。
開始コドンの選択は、リボソームがShine-Dalgarno配列と結合することによって促進されます。 Shine-Dalgarno 配列は、AUG 開始コドンの上流に位置するプリンリッチ領域です。この配列は、16S rRNA のピリミジンリッチ領域に相補的です。 16S rRNA は 30S サブユニットの構成要素です。これらの 2 つの相補的なヌクレオチドが対になって、二本鎖 RNA 構造を形成します。このペアリングは、開始コドンをリボソームの P サイトにもたらします。最初のアミノ酸は P サイトに結合します。リボソームは、A サイト、P サイト、E サイトの 3 つの活性部位で構成されています。最初のアミノアシル tRNA 以外の入ってくるアミノアシル tRNA は、A サイトに結合します。ペプチド結合の形成は P サイトで発生します。非荷電 tRNA の出口部位は E サイトです。
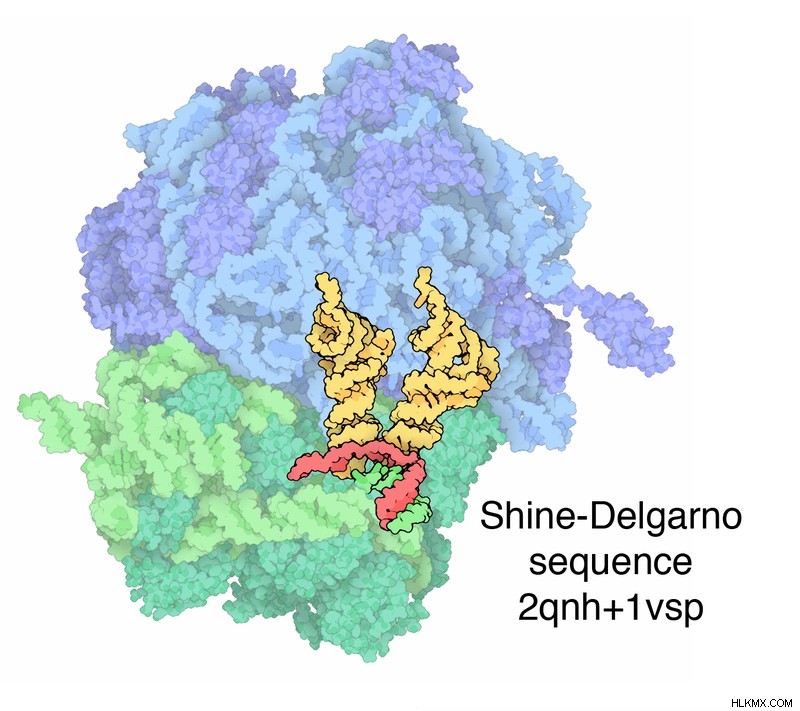
図 1:原核生物の 70S リボソームにおける転写開始
2 つの伸長係数は EF-G と EF-Tu です。翻訳は、リボソームが 3 つの終止コドン (UAA、UGA、UAG) のいずれかに到達するまで延長されます。 tRNA 以外のリリース ファクターは、終止コドンを認識します。 A部位に終結コドンを持つmRNAは、終結複合体と呼ばれます。 RF1、RF2、RF3 の 3 つのリリース要因を特定できます。 RF1 と RF2 は UAA/UAG と UAA/UGA を認識し、ペプチジル tRNA のエステル結合を加水分解して発生期のポリペプチド鎖を放出します。 RF3 は、RF1 と RF2 の放出を触媒します。新しいタンパク質が放出されると、リボソームはリサイクルされます。リボソーム リサイクリング ファクターと EF-G は、リボソームからの mRNA と tRNA の放出、および 70S リボソームの 30S サブユニットと 50S サブユニットへの解離に関与しています。 IF3 は、脱アシル化された tRNA を置き換えることによって mRNA を放出します。
細菌が静止期に入ると、リボソームの二量体化によって翻訳がダウンレギュレートされます。リボソームの二量体化は、RMF、HPF、および YfiA によって促進されます。リボソーム解離因子は RsfA と HflX です。
真核翻訳とは
翻訳は、真核生物の遺伝子発現の第 2 段階であり、真核生物の転写とは別のイベントです。転写と翻訳は、真核生物の 2 つの異なる区画で行われます。したがって、2 つのプロセスが同時に発生することはありません。真核生物のmRNAは単シストロン性であり、細胞質に放出される前に、5 'キャップ、ポリAテールを追加し、イントロンをスプライシングすることにより、核内で処理されます。リボソームの一時停止は、リボソーム上で新たに合成されるポリペプチド鎖の共翻訳フォールディングによって翻訳にも影響を与えます。このプロセスは翻訳を遅らせ、翻訳のための時間を与えます。
真核生物の mRNA は、5' キャップとポリ A テールから構成されます。したがって、翻訳の開始は、キャップ依存性開始とキャップ非依存性開始の 2 つの異なる方法で発生します。キャップ依存性開始の間、開始因子はmRNAの5'末端に結合します。これらの開始因子は、mRNA をリボソームの小サブユニットに保持します。キャップに依存しない開始時に、内部リボソーム エントリ サイトは、直接結合による開始サイトへのリボソームの輸送を可能にします。真核生物では、最初に結合するアミノ酸はメチオニンです。 40S サブユニットは 60S サブユニットと結合して 80S リボソームを形成します。
真核生物の翻訳には、eEF-1 と eEF-2 という 2 つの伸長因子が関与しています。伸長は、原核生物と同様の方法で発生します。翻訳の終結も原核生物系と同じです。しかし、ユニバーサル リリース ファクター eRF1 は、3 つの停止コドンすべてを認識することができます。放出因子である eRF3 は、eRF1 がポリペプチド鎖を放出するのを助けます。翻訳の基本的な手順は 図 2 に示されています。
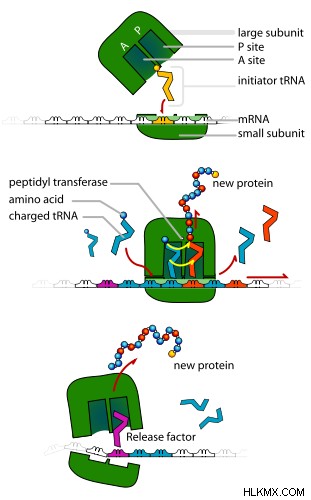
図 2:一般化された翻訳
原核生物と真核生物の翻訳の違い
タイミング
原核翻訳: 原核生物の転写と翻訳は同時進行です。
真核翻訳: 真核生物の転写と翻訳は不連続なプロセスです。
リボソーム
原核翻訳: 30S と 50S =70S リボソーム
真核翻訳: 40S と 60S =80S リボソーム
メッセンジャー RNA ソース
原核翻訳: ポーカリオティック mRNA は細胞質に存在します。 mRNA は多シストロン性です。
真核翻訳: 真核生物のmRNAは核で発生します。転写後修飾の後、それらは核孔を介して細胞質に放出されます。 mRNA は単シストラン性です。
mRNA の寿命
原核翻訳: mRNA は不安定で、数秒から 2 分間存続します。
真核翻訳: mRNA は非常に安定しており、数時間から数日間生存します。
場所
原核翻訳: これは、細胞質の 70S リボソームによって実行されます。
真核翻訳: これは、ER に結合した 80S リボソームによって実行されます。
細胞周期における局在化
原核翻訳: 発生の明確なフェーズはありません。
真核翻訳: これは、細胞周期の G1 期と G2 期で発生します。
未翻訳領域の配列
原核翻訳: Shine-Dalgarno 配列は、開始コドンの 10 ヌクレオチド上流の 5' UTR にあります。
真核翻訳: Kozak 配列は、stat コドンの数ヌクレオチド上流の 5' UTR に見られます。
翻訳開始
原核翻訳: キャップに依存しない開始。
真核翻訳: キャップ依存およびキャップ非依存の両方の開始。
開始要因
原核翻訳: IF1、IF2、IF3 の 3 つの開始因子が関与しています。
真核翻訳: 9 つの開始因子が関与しています:elF 1、2、3、4A、4B、4C、4D、5、および 6。
最初のアミノ酸
原核翻訳: N-ホルミルメチオニンは、ポリペプチド鎖に付加される最初のアミノ酸です。
真核翻訳: メチオニンは、ポリペプチド鎖に付加される最初のアミノ酸です。
伸び係数
原核翻訳: EF-G と EF-Tu の 2 つの伸長因子が関係しています。
真核翻訳: eEF-1 と eEF-2 の 2 つの伸長因子が関与しています。
スピード
原核翻訳: 原核生物の翻訳は、1 秒あたり 20 個のアミノ酸を追加するより高速なプロセスです。
真核翻訳: 真核生物の翻訳は、1 秒あたり 1 つのアミノ酸を追加するより遅いプロセスです。
最初のアミノ酸の運命
原核翻訳: ホルミル基が最初のアミノ酸から除去され、ポリペプチド鎖にメチオニンが保持されます。
真核翻訳: メチオニン全体がポリペプチド鎖から除去されます。
解放要因
原核翻訳: RF1 (UAG と UAA の場合) と RF2 (UAA と UGA の場合) の 2 つのリリース要因が関係します。
真核翻訳: 単一の解放因子が関与します:eRF1.
結論
翻訳は、遺伝子発現の第 2 段階としてタンパク質を合成する普遍的なプロセスです。原核生物と真核生物の両方のリボソームは、基本的に類似した方法で mRNA を解読します。リボソームは、タンパク質合成の機械です。 20 個の必須アミノ酸はすべて、原核生物と真核生物の両方の翻訳プロセスで共有されています。どちらのプロセスも細胞質で発生し、開始、伸長、転座、終結の 4 つのプロセスを完了します。 tRNA は正しいアミノ酸をもたらし、2 つのアミノ酸間でペプチド結合を形成できるようにします。原核生物と真核生物の翻訳の主な違いは、原核生物の翻訳は転写と同時のプロセスであるのに対し、真核生物の翻訳はその転写とは別のプロセスであることです。
参考:
1.「原核翻訳」。ウィキペディア、フリー百科事典、2016 年。2017 年 2 月 26 日にアクセス
2.「真核生物の翻訳」。ウィキペディア、フリー百科事典、2016 年。2017 年 2 月 26 日にアクセス
3.「原核生物と真核生物の翻訳の違い」。 EASY BIOLOGY CLASS、2017。2017 年 2 月 26 日にアクセス
4. Berg JM、Tymoczko JL、Stryer L「真核生物のタンパク質合成は、主に翻訳開始において原核生物のタンパク質合成とは異なります」。生化学。第5版。セクション 29.5、2002 年ニューヨーク:W H フリーマン、ニューヨーク。 NCBIブックシェルフ. 2017 年 2 月 26 日にアクセス
画像提供:
1.「121-70SRibosomes開始」David Goodsell著 – 今月のRCSB PDB Molecule (CC BY 3.0) via Commons Wikimedia
2.「TRNAリボソームダイアグラムen」LadyofHats – Commons Wikimedia経由の自作(パブリックドメイン)