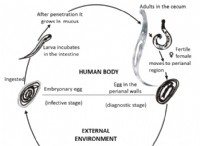
プロテアソーム構造:
-M。結核20Sプロテアソーム:M。tuberculosisの20Sプロテアソームは、2つの積み重ねられた七つの輪で構成され、樽型の構造を形成します。各リングには、7つの同一のβサブユニットが含まれており、中央の触媒チャンバーを形成するために組み立てられています。
- 真核生物26Sプロテアソーム:真核生物26Sプロテアソームはより複雑で、20Sコア粒子と両端の調節粒子で構成されています。 19Sおよび11Sレギュレーターとして知られる調節粒子は、基質認識、展開、および劣化において重要な役割を果たします。
タンパク質分解活性:
-M。結核20Sプロテアソーム:M。tuberculosisの20Sプロテアソームは、エンドペプチダーゼとカルボキシペプチダーゼ活性の両方を示しています。エンドペプチダーゼは、タンパク質基質内のペプチド結合を切断しますが、カルボキシペプチダーゼはC末端からアミノ酸を除去します。
- 真核生物26Sプロテアソーム:真核生物26Sプロテアソームは、主に内ペプチダーゼとして機能し、内部ペプチド結合を切断します。カルボキシペプチダーゼ活性は、通常、26Sプロテアソームに関連していません。
基質特異性:
-M。結核20Sプロテアソーム:結核20Sの基質特異性は完全には理解されていませんが、真核生物プロテアソームとは異なります。結核の20Sプロテアソームは、細胞プロセスに関与するさまざまなタンパク質を分解することができるより広い基質特異性を持っていると考えられています。
- 真核生物26Sプロテアソーム:真核生物26Sプロテアソームは、ユビキチンタグによってマークされた特定の基質を認識および分解します。小さなタンパク質であるユビキチンは、標的タンパク質に共有結合して付着し、26Sプロテアソームによる分解を示します。
細胞調節:
-M。結核20Sプロテアソーム:結核20Sプロテアソームの調節は十分に研究されていません。真核生物26Sプロテアソームで観察される精巧な調節メカニズムがありません。
- 真核生物26Sプロテアソーム:真核生物26Sプロテアソームは、さまざまな細胞シグナルとメカニズムによって厳しく調節されています。ユビキチン化プロセスとその後の調節粒子による認識は、細胞の要求に応じて選択的なタンパク質分解を保証します。
要約すると、結核細胞と真核細胞の両方がタンパク質分解のプロテアソーム系を使用していますが、M。tuberculosisの20Sプロテアソームは、明確な構造的、機能的、調節的特徴を示します。これらの違いを理解することは、結核のプロテアソームを標的とする潜在的な治療戦略を開発するために不可欠であり、最終的に結核との戦いに貢献します。