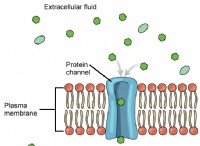
DNAメチル化:DNAメチル化は、遺伝子発現の調節に重要な役割を果たすDNAの化学的修飾です。植物では、精子細胞には、子孫の遺伝子活性に影響を与える可能性のある特定のDNAメチル化パターンが含まれています。これらのパターンは、世代を継承し、その後の世代の遺伝子発現と表現型の特性に影響を与える可能性があります。
ヒストン修飾:ヒストンは、染色体の構造材料であるクロマチンを形成するためにDNAをラップするタンパク質です。アセチル化、メチル化、リン酸化などのヒストンの修飾により、クロマチンの構造が変化し、転写のためのよりアクセスしやすい(エウクロマチン)またはアクセスしやすい(ヘテロクロマチン)。これらの修飾は精子細胞に存在し、子孫の遺伝子発現に影響を与えます。
非コードRNA:非コードRNA(ncrNA)は、タンパク質をコードしないRNA分子です。それらには、マイクロRNA(miRNA)や小さな干渉RNA(siRNA)などの小さなRNA、および長い非コーディングRNA(LNCRNA)が含まれます。精子細胞は、特定のメッセンジャーRNA(mRNA)または調節クロマチン構造を標的とすることにより、転写後に遺伝子発現を調節できるNCRNAを運ぶことができます。
シトシン脱アミノ化:シトシン脱アミノ化は、DNA配列のシトシンをウラシルに変換する化学変化です。これにより、C-to-TまたはG-to-A変異が生じる可能性があります。一部の植物の精子細胞は、高レベルのシトシン脱アミノ化を示し、遺伝的変異に寄与し、子孫の新しい適応につながる可能性があります。
環境の手がかり:母植物が経験する環境の手がかりは、精子を介して子孫に伝染することができます。たとえば、干ばつ、塩分、またはその他の環境ストレスへの曝露は、子孫の遺伝子発現と適応反応に影響を与える可能性のある精子細胞のエピジェネティックな修飾を誘発する可能性があります。
植物の精子の遺伝的配列を超えた情報のエンコードに関する研究は依然として積極的な調査領域であり、将来新しいメカニズムが発見される可能性があることに注意することが重要です。これらのメカニズムは、植物の繁殖と遺伝の複雑さに貢献し、植物が変化する環境条件に適応して対応し、子孫の生存と成功を確保することができます。