科学界は、土壌炭素 (C) と気候の間の正のフィードバック ループを含む気候変動に対する地球の応答を評価するために、地球システム モデル (ESM) の予測に依存しています。温暖化は気候変動をさらに悪化させます。
ただし、土壌従属栄養呼吸、Rh (または、SOM から CO2 への微生物処理) )、二酸化炭素 (CO2) の最大の生態系フラックスの 1 つ )、これらのモデルでは、Q10 として知られる非常に単純化された温度応答関数に基づいています。 .時々このQ10 温度関数には、土壌水分スカラーが伴います。これらの土壌水分スカラーは、多くの場合、非線形 (多項式および放物線) 回帰式を使用して、サイト固有の土壌水分データを Rh に経験的に適合させることによって導出されます。これは、ESM が土壌フラックスの空間的および時間的変動の全範囲を捉えることができないことが多く、これらのモデルの予測可能性を低下させる理由の一部を説明しています。
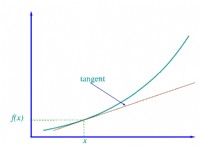
Dual Arrhenius and Michaelis-Menten (DAMM) モデルは、Michaelis-Menten (反応速度が基質との双曲線関係に従う酵素速度論的プロセス)、Arrhenius (指数温度関数)、および拡散 (溶質の輸送と輸送) を使用して Rh をシミュレートします。土壌中の水と空気で満たされた間隙空間を通るガス) 機能 (Davidson et al., 2012, Fig. 1)。
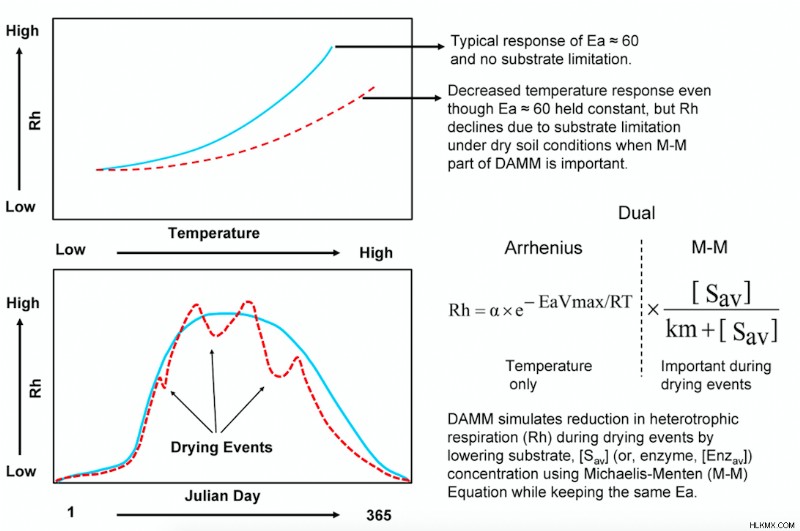
土壌水分は、低水分での可溶性 C 基質の拡散と高水分含有量での酸素の拡散を調節することにより、DAMM の Rh とその見かけの温度感度 (Rh の観測された温度応答) に影響を与えます。最近の研究では、DAMM モデルをより大きな生態系モデルと簡単に統合できることが示されました (Forest Biomass, Assimilation, Allocation and Respiration, FöBAAR; Keenan et al., 2012)、過度に単純化された Q10 を置き換えます 機能し、それによって生態系モデルのパフォーマンスを向上させます (図 2)。そうすることで、他のエコシステム モデルや ESM がどのように彼らのアプローチを採用できるかを示すつもりです。
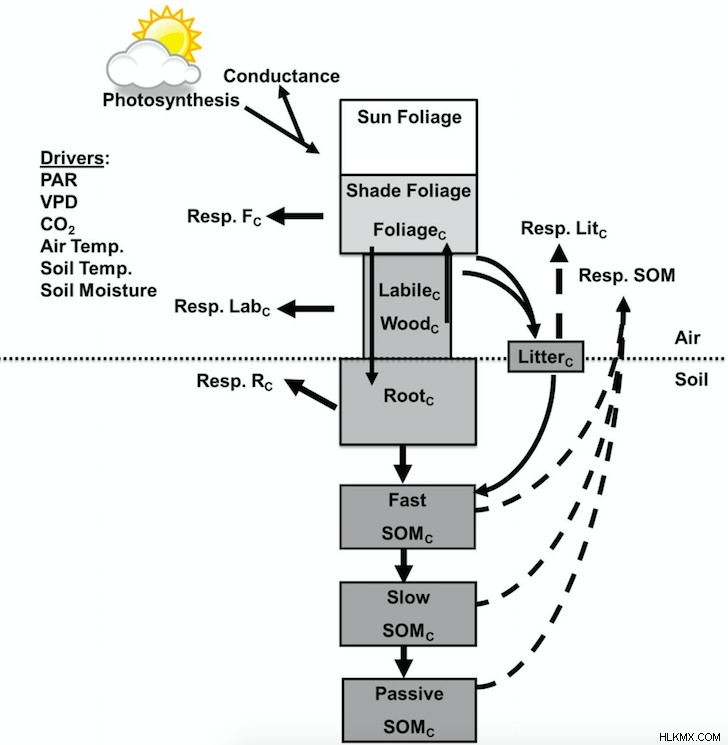
シヒら。 (2018) 米国北東部の 2 つの AmeriFlux サイトで、同一のデータセット (自動土壌チャンバーからの高周波土壌フラックス データ、渦共分散タワーからのランドスケープ スケールの生態系フラックス、および落葉や葉面積指数などの他のキャンペーン ベースの測定値) を使用しました (マサチューセッツ州ハーバード フォレストおよびメイン州ハウランド フォレスト) は、反復プロセスを使用して、マージされた (DAMM-FöBAAR) モデルとマージされていない (FöBAAR のみ) モデルを調整します (Williams et al., 2009)。根排除 (トレンチング) 実験からの土壌フラックスを使用して、DAMM-FöBAAR モデルの Rh コンポーネントを直接パラメータ化しました。次に、DAMM-FöBAAR モデルと FöBAAR のみのモデルの両方のパフォーマンスが、短期 (季節) および長期 (10 年) の土壌 (土壌微生物と根の呼吸の合計) および生態系 (独立栄養呼吸と従属栄養呼吸の合計) フラックスについて検証されました。 .キャリブレーションされた DAMM-FöBAAR モデルは、両方のサイトの FöBAAR のみのモデルと比較して、季節的および 10 年規模の土壌 (および生態系) 呼吸に関するモデル データの不一致を低減し、極端な土壌水分変動 (乾燥など) がある条件下でのパフォーマンスが大幅に向上しました。イベント)
上記の調査結果は、DAMM-FöBAAR モデルにおける Rh の土壌水分 (および基質供給) 効果のより明確な表現がロバストであることを示しています。 時間と空間を超えて。さらに重要なことに、DAMM モデルのミカエリス-メンテン方程式は、温度感度パラメーターを一定に保ちながら、乾燥 (およびフラッディング下の酸素) 条件下での基質-C 制限を制御します (つまり、アレニウス関数の「Ea」、図 1)。 .これは、従来の Q10 を搭載したモデルよりも大きな利点です。 Q10 のような表現 に基づくモデルは、同様の環境条件下で調整されていない限り、乾燥 (または洪水) イベント下での土壌呼吸を再現することはできません。したがって、Sihi et al。 (2018) は、DAMM モデル方程式が土壌 (および生態系) 呼吸の予測能力を増加させることを実証しましたが、モデルの複雑さは比較的わずかです。
最後に、DAMM モデルの方程式における Rh の基質供給に対する土壌水分の影響のシンプルかつ明確な表現は、一般的に使用されるさまざまな経験的回帰方程式よりも一般化可能であり、より大きな ESM に容易に組み込むことができます。
これらの調査結果は、米国北東部の 2 つの AmeriFlux サイトにおける土壌従属栄養呼吸の機械的酵素モデルを生態系モデルに統合するというタイトルの記事で説明されています。
この作業は、メリーランド大学環境科学センターの Debjani Sihi (現在 ORNL に在籍) と Eric A. Davidson、パシフィック ノースウェスト国立研究所の Min Chen、ウッズホール研究センターの Kathleen E. Savage、Andrew G.ハーバード大学と北アリゾナ大学の Richardson、ローレンス バークレー国立研究所の Trevor F. Keenan、および USDA Forest Service の David Y. Hollinger。
参考文献:
<オール>