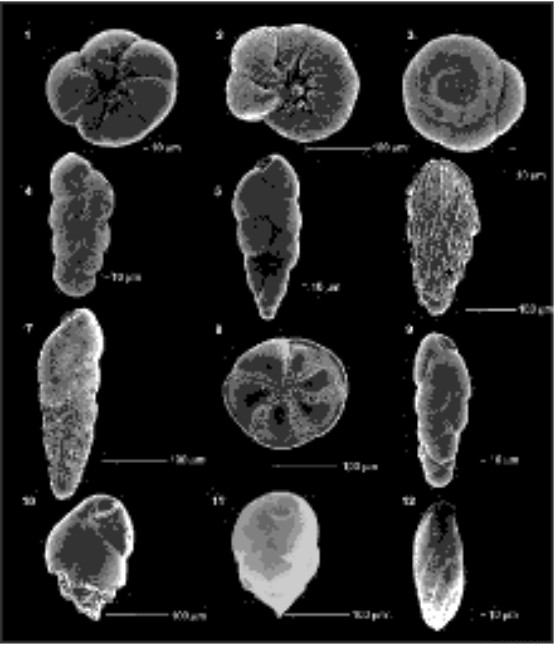
教育と研究に対する私の魅力は、教授とポスドク時代の 15 年間の大学院での教育と指導によって培われてきました。私の教職歴には、環境指標と多変量解析を使用して、底生有孔虫種 (図 1) と堆積物と水との境界面における非生物的パラメーターとの間の相互作用を空間的および時間的スケールで理解するための議論、フィールド サンプリング、実験手順が含まれていました。
私は生物海洋学者であり、生態学、堆積学、海洋地質学、生物地球化学、堆積学、地球生物科学、古気候学などのさまざまな分野に興味を持っていますが、これらに限定されません。特に有孔虫の種パターンに興味があります。これは堆積学的特性、気候変動、現在のマングローブ、河口、サンゴ礁、大陸棚、深層水の汚染に関連する最近の海と淡水の混合を反映しています。
私は、IODP Leg 363 で回収された場所を使用して、底生有孔虫群集が西太平洋の底水温と栄養レベルに敏感であるかどうかを調べるのが好きです。私は温暖水種と寒冷種に適応した種を研究し、温水種曲線 (温水種の割合) を確立します。ウォーム プラス コールド 適応フォームの合計に関して)。また、空間スケールで、最も若い間隔 (最初のコアのコア キャッチャー) からの温水種の割合が、現代の底層水温と有意に相関している可能性があることを示すためにも取り組んでいます。これは、底生有孔虫が温度に敏感であることを裏付けており、研究サイトによって囲まれた狭い範囲の温度 (~2-5 ℃) に対してさえもです。ダウンコアの変動は、より一般的な意味ではありますが、地球規模の気候背景条件に関連している可能性があります。私の研究は、底水温度のトレーサーとして底生有孔虫群集の使用を支持し、フォローアップのより高解像度の研究の正当性を提供します.
また、海洋貯留層からのメタンの放出は、第四紀の気温変化による気候変動と関連しています。この仮定は、私が研究している群集の動物相と地球化学に対するメタンフラックスの応答による、底生有孔虫の安定同位体サインの変動に基づいています。 Uvigerina としてのコスモポリタンな石灰質有孔虫群集 、ボリビア 、キロストメラ 、グロブリミナ 、およびノニオネラ 有機物が豊富な環境での冷たいメタン浸透からの浸透に関連する、古海洋学的に重要な分類群の一部でした。
湧き出る有孔虫は、冷たい湧出での食物の入手可能性に惹かれ、有機物が豊富で還元的な環境での生活に必要なものを超えて適応する必要はありません。有孔虫試験の d13C 値は、メタンの浸出と同位体組成の種固有の違いを反映しており、浸出活動の時間的変動を示している可能性があることを示唆しています。熱水変質と動員は、埋もれた炭素を水圏と潜在的に大気に再注入します。このプロセスは、有孔虫によって追跡できる地球の歴史を通じて、地球規模の気候と大量絶滅イベントの主要な変化に関連していると考えられています。地下の微生物集団は、これらの熱水で生成され動員された炭素源、炭化水素、およびメタンを遮断して処理することができます。
有機物に富む堆積物、熱変化、および関連する炭素放出に関連する大量のマグマは、堆積物と水の界面に住む動物相によって検出されます。したがって、有孔虫の生態と安定同位体組成をよりよく理解することで、古湧水の認識が向上し、気候と古海洋学的変化の解釈が改善されます。有孔虫の個体群動態とそれらの相互作用に焦点を当て、過去の堆積物の変化と蓄積と土台の定置を指摘しようとしています.
海洋領域からのものなどの高解像度の連続記録は、大量絶滅の考えられる原因を調べるために、化石の発生を地球化学的環境プロキシと組み合わせる素晴らしい機会を提供します。たとえば、更新世中期の遷移 (MPT、約 1.2 ~ 0.6 Ma) は、地球規模の冷却、氷河期の延長、海洋循環の変化、および陸生生物と海洋生物相の進化によって特徴付けられました。 MPT の間、100 種を超える深海の底生有孔虫が段階的に絶滅し、特定の形態学的グループ (主に「絶滅グループ」として知られる、装飾された開口部を持つ細長い「stilostomellids」) を標的にしました。水深 1600 ~ 2000 m の冷たいメタンの上部コアからの生きている (ローズ ベンガル染色) およびコアからの死んだ (化石) 有孔虫 (>150mm) の有孔虫の生態と安定同位体組成。過去と現在のメタン放出の再構築の潜在的な用途を調査するために、堆積物の変動を捉えた低解像度の記録を生成し、これらのメタンの浸透で深部生物圏の生命を制限する条件の理解を深めます。