エピジェネティクスとは、遺伝子の発現の遺伝的変化、または遺伝子のヌクレオチド配列の変化によって生じない特定の生物の表現型の遺伝的変化に関する研究です。 .遺伝子発現のエピジェネティックな調節は、組織特異的な遺伝子発現、X 染色体の不活性化、およびゲノム刷り込み (親固有の方法での遺伝子の発現) に関与するため、細胞機能において重要な役割を果たします。さらに、エピジェネティックに調節される遺伝子の発現障害は、がんなどの疾患を引き起こします。エピジェネティックな遺伝子調節に関与するメカニズムは、DNA メチル化、非翻訳 RNA、クロマチン構造、および修飾です。この記事では、遺伝子発現に対する DNA メチル化の影響について説明します。
対象となる主な分野
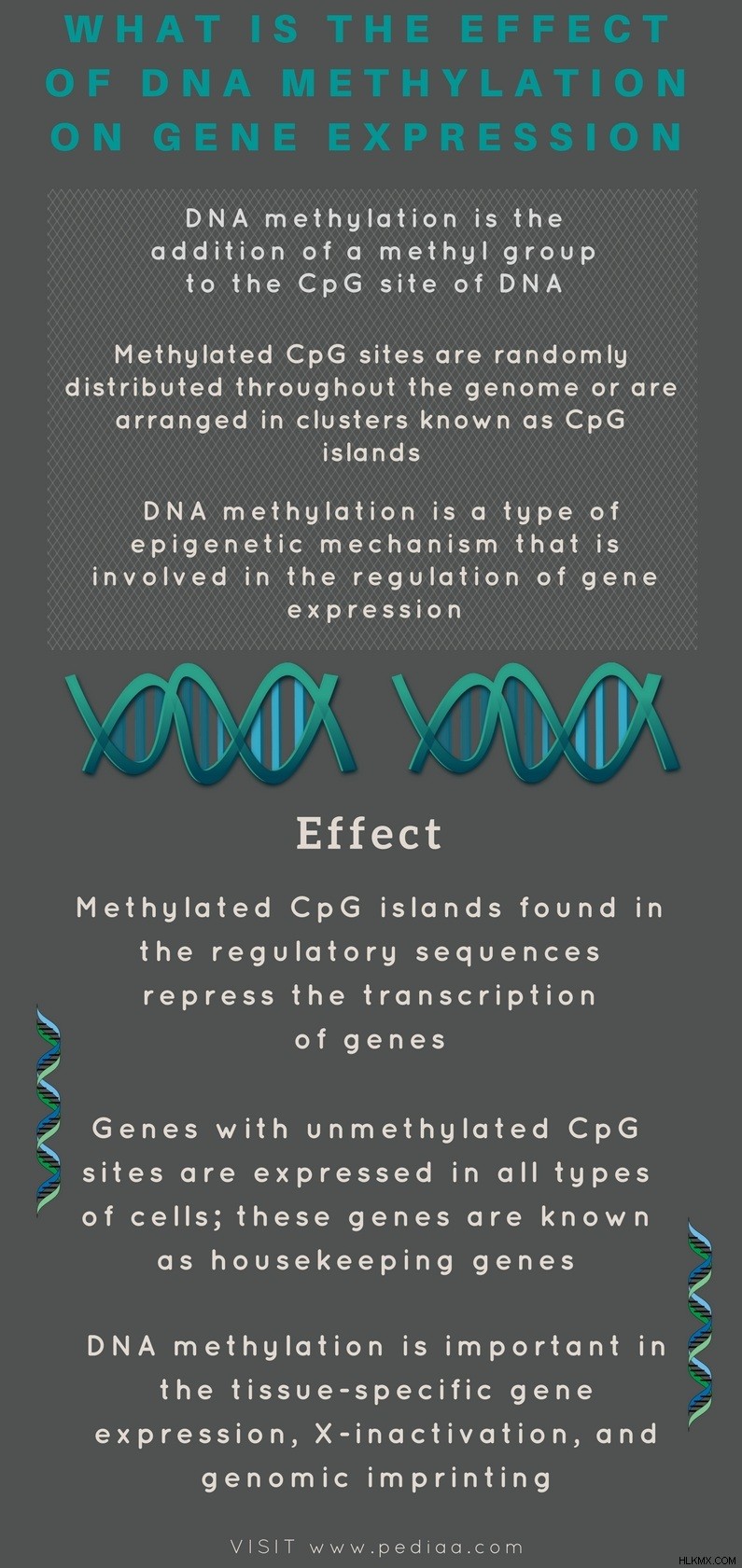
1. DNAメチル化とは
– 定義、ゲノム内分布、重要性
2.遺伝子発現に対する DNA メチル化の影響
– メチル化の機能
3.細胞機能における DNA メチル化の役割
– 組織特異的遺伝子発現、X 染色体の不活性化、ゲノム刷り込み
重要な用語:CpG アイランド、DNA メチル化、エピジェネティクス、ゲノム刷り込み、組織特異的遺伝子発現、X 不活化
DNA メチル化とは
DNA メチル化とは、メチル基 (-CH3) の付加を指します ) 5'-CpG-3' 部位で窒素ベースのシトシン (C) に共有結合。 CpG部位は、直鎖DNA鎖の5'から3'方向に沿ってシトシンヌクレオチドの後にグアニンヌクレオチドが続くDNAの領域である。シトシンは、リン酸(p)基を介してグアニンヌクレオチドに結合しています。 DNA メチル化は、DNA メチルトランスフェラーゼによって調節されます。メチル化されていないシトシンとメチル化されたシトシンを 図 1 に示します .

図 1:非メチル化およびメチル化シトシン
メチル化されていない CpG サイトは、ランダムに分散するか、クラスターに配置することができます。 CpG サイトのクラスターは「CpG アイランド」と呼ばれます。これらの CpG アイランドは、多くの遺伝子のプロモーター領域に存在します。ほとんどの細胞で発現するハウスキーピング遺伝子には、メチル化されていない CpG アイランドが含まれています。多くの場合、メチル化された CpG アイランドは遺伝子の抑制を引き起こします。したがって、DNAメチル化は、さまざまな組織における遺伝子の発現を制御するだけでなく、胚発生などの特定の時期に制御します。進化を通じて、複製された転移因子、反復配列、およびウイルス DNA などの外来 DNA をサイレンシングする際の宿主細胞の防御メカニズムとして、DNA メチル化は重要です。
遺伝子発現に対する DNA メチル化の影響とは
ゲノムの CpG サイトのエピジェネティックなマーキングは、種に固有のものです。それは遺伝するだけでなく、生涯を通じて安定しています。ヒトゲノムでは、多くの CpG サイトがメチル化されています。 DNA メチル化の主な機能は、特定の細胞の要件に応じて遺伝子発現を調節することです。 図 2 は、哺乳類における典型的な DNA メチル化の状況を示しています。 .

図 2:哺乳類における DNA メチル化の状況
遺伝子発現は、エンハンサーなどの遺伝子の調節配列に転写因子が結合することによって開始されます。 DNAメチル化によってクロマチン構造にもたらされる変化は、転写因子の調節配列へのアクセスを制限します。さらに、メチル化された CpG サイトは、メチル CpG 結合ドメイン タンパク質を引き付け、ヒストン修飾の原因となるリプレッサー複合体を動員します。ヒストンは、DNA のラッピングを変更するクロマチンのタンパク質成分です。これにより、ヘテロクロマチンとして知られるより凝縮されたクロマチン構造が形成され、遺伝子発現が阻害されます。それどころか、ユークロマチンは、遺伝子発現を可能にするゆるいクロマチン構造の一種です。
細胞機能における DNA メチル化の役割とは
一般に、特定の細胞の DNA メチル化パターンは非常に安定しており、特異的です。組織特異的な遺伝子発現、X 染色体の不活性化、およびゲノム刷り込みに関与しています。
組織特異的遺伝子発現
組織の細胞は、体内で特定の機能を実行するために分化しています。したがって、細胞の構造的、機能的、および調節要素として機能するタンパク質は、異なる方法で発現する必要があります。このタンパク質の異なる発現は、各タイプの組織における遺伝子の DNA メチル化の異なるパターンによって達成されます。特定の生物のあらゆる種類の細胞のゲノムの遺伝子は同じであるため、組織で発現する必要のない遺伝子は、その調節配列にメチル化された CpG アイランドを含んでいます。ただし、胚発生中の DNA メチル化のパターンは成体段階のものとは異なります。がん細胞では、DNA メチル化の規則的なパターンは、その組織の正常な細胞とは異なります。正常細胞と癌細胞の DNA メチル化パターンを図 3に示します。 .
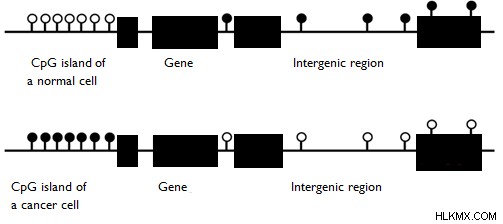
図 3:正常細胞と癌細胞の DNA メチル化パターン
X 染色体の不活化
女性は 2 つの X 染色体を持ち、男性はゲノムに X 染色体と Y 染色体を持っています。女性の X 染色体の 1 つが発生中に不活性化されます。これは、de novo メチル化によって達成されます。 X染色体の不活性化は、ヘテロクロマチンを形成することにより、サイレント段階に維持されます。 X不活性化は、X染色体に関連する遺伝子産物の発現を男性の2倍防ぎます。胎盤哺乳類では、X 染色体を不活性化する選択はランダムです。ただし、不活性化すると、生涯無音のままです。しかし、有袋類では、父方由来の X 染色体のみが不活性化されています。
ゲノム刷り込み
ゲノム刷り込みとは、親染色体の起源に応じて遺伝子を選択的に発現させることです。例として、インスリン様成長因子 2 (IGF2) の父親のコピー ) 遺伝子はアクティブですが、母体のコピーはアクティブではありません。ただし、H19 の場合は逆です。 IGF2 に密接に位置する遺伝子 遺伝子は同じ染色体にあります。ヒトゲノムの約 80 の遺伝子が刷り込まれています。 DNA メチル化は、特定の遺伝子の 1 つの親コピーの不活化に関与しています。
結論
遺伝子のエピジェネティックな変化による遺伝子発現の調節は、多くのゲノムの安定した遺伝特性です。エピジェネティックな遺伝子調節の重要なメカニズムの 1 つは、DNA メチル化です。 DNA メチル化とは、CpG サイトのシトシン残基にメチル基を永続的に付加することです。遺伝子の調節配列の近くにあるメチル化された CpG アイランドは、その特定の遺伝子の転写を抑制します。したがって、これらの遺伝子は沈黙したままです。 DNA メチル化による遺伝子のサイレンシングは、組織特異的な遺伝子発現、X 不活性化、およびゲノム刷り込みにおいて重要です。
参照:
1. リム、デレク H K、イーモン R マハー。 「DNAメチル化:遺伝子発現の後成的制御の一形態」 The Obstetrician &Gynaecologist、Blackwell Publishing Ltd、2011 年 1 月 24 日、こちらから入手可能。
2. ラジン、A、H シダー。 「DNAメチル化と遺伝子発現。」 Microbiological reviews.、米国国立医学図書館、1991 年 9 月、こちらから入手可能。
画像提供:
1. マリウスウォルターによる「DNA メチル化」 – 自作 (CC BY-SA 4.0) コモンズ ウィキメディア経由
2. マリウスウォルターによる「DNAme ランドスケープ」 – 自作 (CC BY-SA 4.0) コモンズ ウィキメディア経由
3.「正常細胞とがん細胞の DNA メチル化」Ssridhar17 著 – Commons Wikimedia による自作 (CC BY-SA 4.0)