コア コンセプト
この生化学のチュートリアルでは、ワトソンとクリックの塩基対について、その重要性、構造、化学、およびいくつかの重要な非ワトソン-クリック塩基対について学びます。
DNA のワトソン クリック モデルとは?
1953 年、生化学者のジェームズ ワトソンとフランシス クリックは、DNA 二重らせんの最初のモデルを発表しました。彼らは、モーリス・ウィルキンスとロザリンド・フランクリンが X 線結晶構造解析を使用して行った DNA イメージングを通じて、この今や有名なモデルを開発することができました。 DNA 構造の Watson-Crick モデルは、最も一般的な DNA 互変異性体である B-DNA に特異的に対応し、多くの正確な詳細が含まれていました。
- リン酸塩と糖は外側を向いていますが、窒素塩基はらせんの内側で対になったままです。その結果、「ねじれたはしご」ができます
- ストランドの逆平行方向 (3'-5' および 5'-3')
- 主溝と副溝の存在
- DNA の右回りの時計回りのねじれ
このモデルの最も画期的な詳細は、適切にワトソン-クリック塩基対と名付けられた窒素塩基の対形成でした。 DNA 生化学の主要な要素と考えられているワトソン-クリック塩基対形成には、アデニンとチミンの対形成、およびグアニンとシトシンの対形成が含まれます。または、生物学の学生がよく覚えているように、A は T に、G は C に。
この功績により、ワトソンとクリックは 1962 年のノーベル生理学・医学賞をモーリス ウィルキンズと共同受賞しました。ロザリンド・フランクリンは、彼女の貢献に対して 1962 年の賞を授与されませんでした。
窒素塩基の構造 ワトソンクリック塩基対で
DNA の 4 つの窒素塩基は、次の 2 つのグループに分けることができます。
- ピリミジン:チミンとシトシン
- プリン:アデニンとグアニン
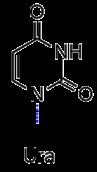
有機化学では、ピリミジンは 1 位と 3 位に 2 つの窒素を持つベンゼンのような芳香環です。ピリミジンの誘導体として、チミン、シトシンは環構造に結合した固有の置換基を持っています。シトシンは炭素 4 に結合したアミノ基と炭素 2 にカルボニルを持っています。逆に、チミンは炭素 2 と 4 に 2 つのカルボニルと炭素 5 にメチル基を持っています。チミンの RNA 等価物であるウラシルもピリミジンとしてカウントされます。 、チミンからメチルを引いたものと同じ構造です。
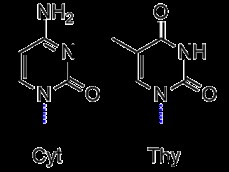
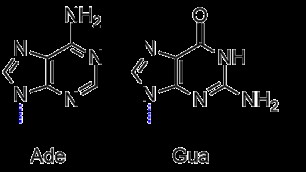
対照的に、プリンは、1、3、7、および 9 位に 4 つの窒素ヘテロ原子を含む 9 員環の芳香族二重環です。アデニンとグアニンは、この基本構造を維持しますが、特徴的な置換基を持っています。アデニンには、炭素 6 に結合したアミノ基があります。しかし、グアニンには、炭素 2 に結合したアミノ基と、炭素 6 にカルボニルを結合したものがあります。
彼らのモデルのこの要素について、ワトソンとクリックは、DNA のどのストレッチにおいても、プリン (アデニンとグアニン) の合計が常にピリミジン (チミンとシトシン) の合計に等しいというアーウィン チャーガフの最近の発見の形で重要なヒントを得ました。このことから、ワトソンとクリックは、両方のプリンが対応するピリミジンを持っていることを発見しました.
ワトソン-クリック塩基対:アデニンとチミン
ワトソンとクリックは、アデニンとチミンには、水素結合を同時に形成できる 2 対の化学基があることを観察しました。
- アデニンのアミノとシトシンのカルボニル
- アデニンの六員環の不飽和窒素とチミンの環の飽和窒素
ワトソン-クリック塩基対:グアニンとシトシン
ワトソンとクリックは、グアニンとシトシンが同時に水素結合を形成できる 3 対の化学基を持っていることを観察しました:
- グアニンのカルボニルとシトシンのアミノ
- グアニンの六員環の飽和窒素とシトシンの環の不飽和窒素
- グアニンのアミノとシトシンのカルボニル

さらに、グアニン-シトシンのペアは、アデニン-チミンのペアよりも水素結合が 1 つ多いため、DNA 鎖は、グアニン-シトシンのペアが多いほど、または「GC」含有量が多いほど、相互に強い親和性を持ちます。 GC 含有量が高くなると、DNA 鎖を自然に分離するのに必要な「融解」または「変性」温度が上昇します。これは、特定の温度での DNA 鎖の分離に依存する PCR などの特定の分子生物学ラボ技術に重要な意味を持ちます。
非ワトソン クリック塩基対
DNA や RNA もワトソン クリックの塩基対のみを含む傾向がありますが、「標準的でない」塩基対がさらに存在します。これらの代替塩基対は、酸性条件下または特定の RNA 構造の下で DNA に見られます。
フーグスティーン塩基対
Hoogsteen 塩基対では、グアニンは依然としてシトシンと対になり、アデニンは依然としてチミンと対になります。ただし、ピリミジン塩基、グアニンおよびアデニンは、ワトソンクリック方向に対して 180°「反転」します。生化学者は、ワトソン-クリックの塩基対形成におけるアンチ方向とは対照的に、この「反転した」方向をシン確認と呼んでいます。
アデニンとチミンの間では、次のグループのペアの間に 2 つの水素結合が形成されます:
- アデニンのアミノ基とチミンのカルボニル
- アデニンの 5 員環の不飽和窒素とチミンの環の飽和窒素
グアニンとシトシンの間では、次のグループのペアの間に (3 ではなく) 2 つの水素結合が形成されます:
- グアニンのカルボニルとシトシンのアミノ
- グアニンの 5 員環の不飽和窒素とチミンの環のプロトン化窒素
注:グアニン-シトシン ペアでは、必要な水素結合を形成するために、シトシンの環窒素をプロトン化する必要があります。このため、Hoogsteen 塩基対形成は低 pH で優先される傾向があり、一方 Watson-Crick 塩基対形成は中性または高 pH で優勢です。
興味深いことに、アデニンとグアニンの通常の結合基は対になっていないため、Hoogsteen 塩基対によって三重鎖 DNA が可能です。ただし、これらの構造はほとんどの条件下では形成されない傾向があります。
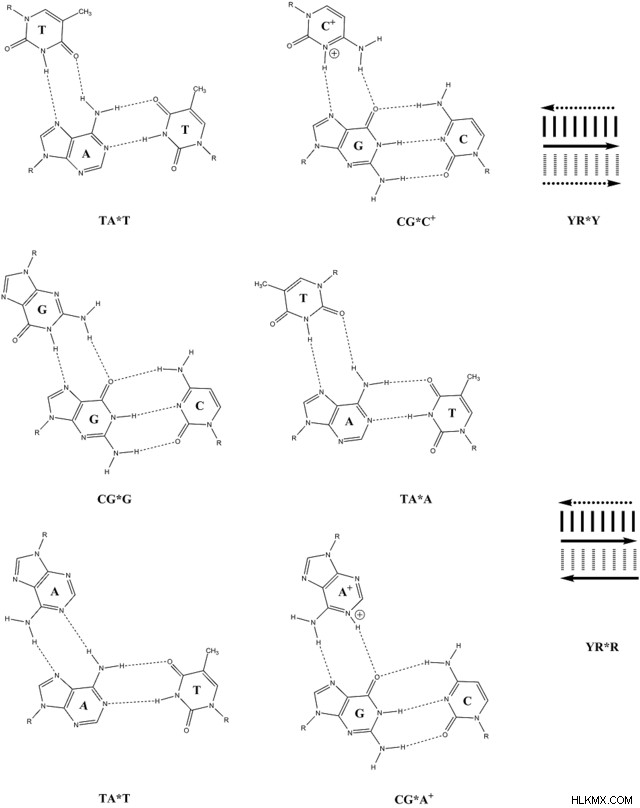
ウォブル ベース ペア
転写では、RNAポリメラーゼはワトソン-クリック塩基対合に依存してmRNA鎖のコードを決定します。しかし、翻訳では、生化学者が「ゆらぎ」塩基対と呼ぶものも、mRNA と tRNA の間の相互作用に存在します。
興味深いことに、tRNA には、ヒポキサンチンと呼ばれる別のピリミジン塩基があり、これがヌクレオチド イノシンを形成します。

ヒポキサンチンは、アミノ基を除いたグアニンに似ています。機能的には、ヒポキサンチンは、ウラシル、シトシン、およびアデニンと塩基対で 2 つの水素結合を形成します。これらの水素結合は、ヒポキサンチンのカルボニルと飽和窒素から特異的に形成されます。
ヒポキサンチンとのこれらの 3 つの新しい塩基対の他に、グアニンはウラシルとの不安定な塩基対を形成することもできます。両方の水素結合は、カルボニルと飽和窒素の間に形成されます。
これらの代替の非ワトソンクリック塩基対により、遺伝コードの冗長性が可能になります。 tRNA アンチコドンの 3 番目の位置に配置されると、ウラシル、グアニン、およびヒポキサンチンは複数の可能な mRNA 塩基と結合できます。
たとえば、UAI のアンチコドンを持つアミノアシル tRNA は、常にイソロイシン アミノ酸を持っています。ヒポキサンチンの不安定な塩基対形成により、AUC、AUA、および AUU の mRNA コドンがすべてヒポキサンチンに結合し、したがってすべて最終タンパク質のイソロイシンをコードします。