コア コンセプト
この生化学の記事では、生物学のセントラル ドグマについて、そのさまざまな定義、ドグマを一般的にカバーする細胞プロセス、およびいくつかの特殊なケースを含めて学びます。
生物学のセントラル ドグマとは?
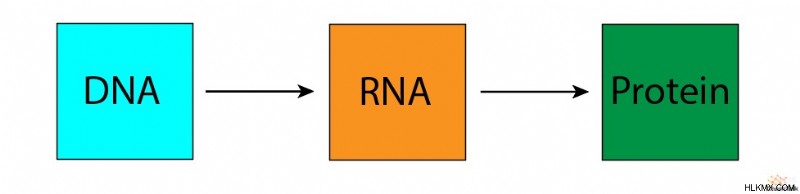
生物学のセントラル ドグマでは、生物は DNA から RNA、RNA からタンパク質への生物学的情報の流れに基づいて機能すると述べています。
生物は複雑です。生物を構築するには、その生化学的構成要素を合成し、非常に具体的なガイドラインに従って使用する必要があります。これらのガイドラインは DNA の形で提供されます。しかし、DNA はそれ自体で生物の生物学的機能を実行することはできません。代わりに、タンパク質は、その構造の生化学的特性に従って、これらすべての機能を実行します。重要なことは、DNA の生物学的情報がこれらのタンパク質構造を特定することです。 RNA は、DNA の遺伝情報をタンパク質の構造に伝達するメッセンジャーとして機能します。 DNA から RNA を経てタンパク質に至るこの情報の流れは、分子生物学のほとんどすべての理解が依存する重要な基礎を提供します。その結果、科学者はこれを生物学のセントラル ドグマと呼んでいます。
生物学のセントラル ドグマは一方向ですか?
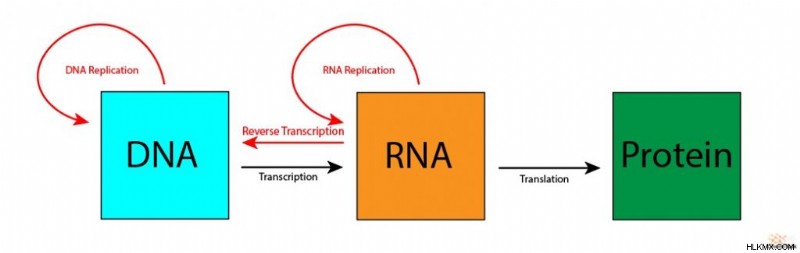
多くの文脈で、セントラル ドグマは、DNA から RNA への転写による生物学的情報の「一方向」の流れ、RNA から翻訳によるタンパク質への生物学的情報の流れとして説明されています。この解釈は生物学的情報の最も重要な流れをカバーしていますが、DNA 複製などの多くの情報伝達は除外されています。
セントラル ドグマのより正確なバージョンは、元の意味で見つけることができます。 DNA のらせん構造の共同発見者の 1 人であるフランシス クリックは、1957 年に初めてセントラル ドグマを発表しました。彼は、情報は核酸 (DNA と RNA) から核酸へ、または核酸からタンパク質へのみ伝達されると述べました。しかし、情報はタンパク質からタンパク質へ、またはタンパク質から核酸へと流れません。
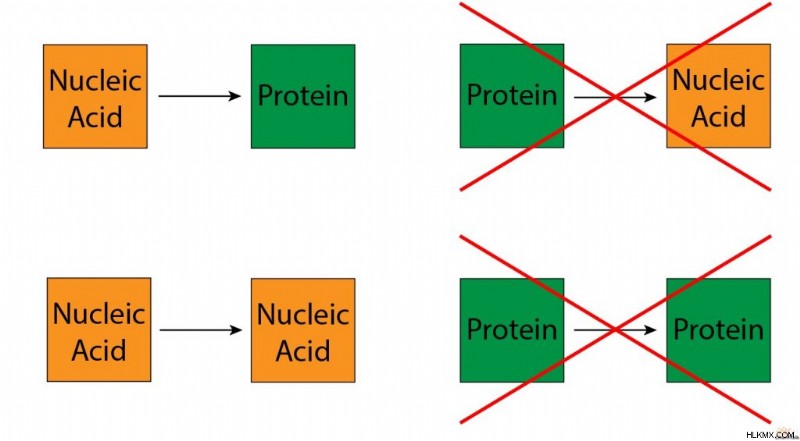
Crick のバージョンには、転写、翻訳、DNA 複製などの従来の生物学的情報の転送が組み込まれています。さらに、逆転写や RNA 複製など、存在することが証明されているが、「一方向」の流れに反するいくつかの「特別な転送」が含まれています。
セントラル ドグマ ステップ 1:DNA から RNA へ
機能的な生物学的システムになるには、まず情報が DNA から RNA に転送されなければなりません。細胞は、転写と呼ばれるプロセスを通じてこれを行います。文字起こしに関する詳細な説明については、こちらの記事をご覧ください。
最終的にタンパク質の構造を知らせる DNA に記録された情報は、遺伝子の形で提供されます。これらの遺伝子は、アデニン、グアニン、シトシン、およびチミン (AGCT) の 4 つの窒素塩基のいずれかを持つヌクレオチドの配列です。驚くほど単純ですが、これら 4 つのヌクレオチドの配列は、タンパク質を構築するために必要なすべての情報を提供します。
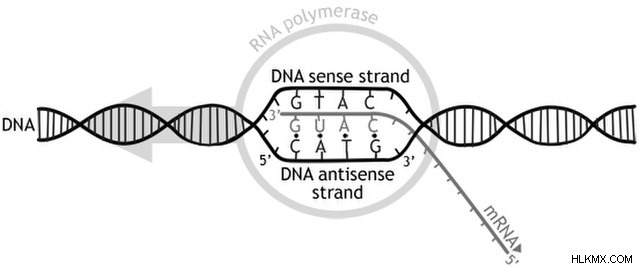
DNA がタンパク質の合成を指示できるようになる前に、その配列はまず RNA に転写されなければなりません。ワトソン・クリックの塩基対合規則 (A と T、G と C) を使用して、酵素 RNA ポリメラーゼは DNA の配列に沿って RNA 鎖を構築します。得られたmRNA鎖、または「メッセンジャー」RNAは、DNA遺伝子に相補的な配列を持っています。したがって、RNA 転写を通じて、DNA の生物学的情報が RNA に転送され、RNA は核を離れてタンパク質を合成することができます。
セントラル ドグマ ステップ 2:RNA からタンパク質へ
生物学的情報の利用を完了するには、転写された RNA 遺伝子がタンパク質の構造に変換されなければなりません。これは、翻訳と呼ばれるプロセスを通じて行われます。翻訳に関するより詳しい説明については、こちらの記事をご覧ください。
具体的には、核内で合成された mRNA 鎖が、タンパク質のアミノ酸配列を知らせます。これを行うために、鎖はタンパク質合成を担う細胞小器官であるリボソームと相互作用します。中間体として tRNA の分子を使用して、リボソームは「コドン」と呼ばれる 3 つの mRNA 塩基のグループを「読み取り」ます。このグループは、20 の一般的なアミノ酸の 1 つをコードします。
したがって、それ自体が DNA 配列に基づいている mRNA の配列に基づいて、リボソームは一連のアミノ酸をつなぎ合わせます。この配列は、タンパク質の最も基本的な、または「一次」構造を提供します。
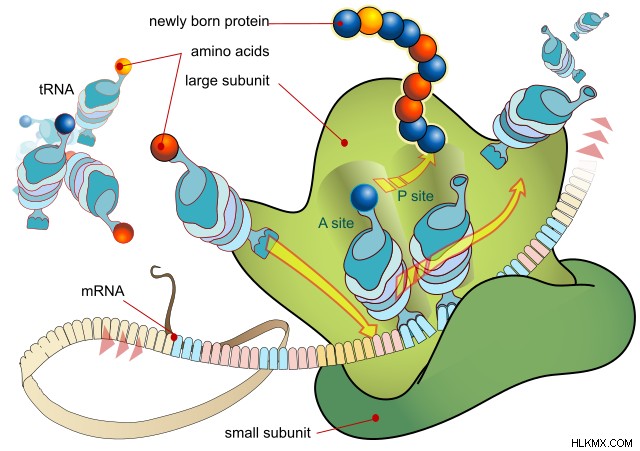
合成されると、アミノ酸配列はゴルジ体で機能性タンパク質に折り畳まれます。重要なのは、特定のアミノ酸グループが互いに自然に相互作用するため、アミノ酸の順序によってタンパク質が最終的にどのように折り畳まれるかが決まる.その後、折り畳まれたタンパク質は生体内で必要な機能を引き受けます。
セントラル ドグマの特殊なケース
セントラル ドグマの「一方向」バージョンを構成する転写と翻訳以外にも、生物学的情報の多くの伝達が行われます。ただし、これらの「特殊なケース」にはそれぞれ、DNA 以外の核酸から RNA への転移が含まれます。
DNA から DNA:DNA 複製
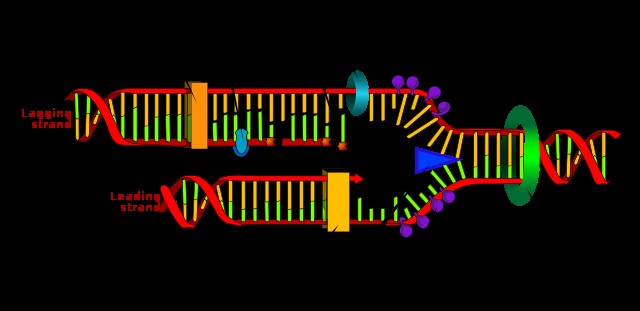
前述のように、「一方向」の解釈に含まれない最も一般的な情報伝達の形式は DNA 複製です。このプロセスでは、1 つの DNA 二重らせんの情報が 2 つの半保存された DNA らせんに転送されます。 DNA 複製について詳しくは、この記事をご覧ください。
転写と同様に、DNA 複製は相補的な塩基対に依存します。このプロセスには、ポリメラーゼ、リガーゼ、プライマーゼ、ジャイレース、トポイソメラーゼなど、さまざまなタンパク質が必要です。本質的に、DNA 二重らせんが分離され、2 つの新しい DNA 鎖が親鎖の塩基対合によって合成されます。これにより、最終的に 1 つのオリジナルから 2 つの完全な二重らせんが得られます。
RNA から RNA:RNA 複製
DNA 複製ほど一般的ではなく、よく知られている RNA 複製も、あるタイプの核酸から同じタイプの新しい鎖への情報の転送を含みます。今回は RNA を使用します。
DNA の複製や転写と同様に、RNA の複製にも新しい鎖を合成するための相補的な塩基対形成が含まれます。このプロセスには、RNA 依存性 RNA ポリメラーゼ (RdRp) と呼ばれる酵素が必要です。
真核生物は主に RdRp を使用して、RNA 干渉のためのガイド RNA 鎖を合成します。これらの新しい相補的 RNA 鎖は、特定の配列の mRNA に結合し、二本鎖 RNA (dsRNA) を形成します。次に、タンパク質 Dicer が dsRNA に結合し、鎖を切断して短い翻訳不可能な配列にします。 RNA 干渉により、真核生物は特定の遺伝子のタンパク質への翻訳を阻害することで、その遺伝子の発現を変えることができます。
RdRp による RNA 複製は、レトロ ウイルスに感染した細胞でも発生します。 RdRp 遺伝子は、レトロウイルス RNA プラスミド上に存在することがよくあります。感染細胞でのRdRpの産生により、DNAを必要とせずにRNAプラスミドの複製と増幅が可能になるため、これはウイルスに役立ちます.
RNA から DNA:逆転写
セントラル ドグマの「一方向性」モデルと完全に矛盾しており、RNA は特定の状況で DNA への逆転写を示しています。
繰り返しになりますが、RNA と DNA の両方の転写と複製と同様に、逆転写には相補的な塩基対形成が含まれます。このプロセスには逆転写酵素 (RT) という酵素が必要です。 RdRp と同様に、RT 遺伝子はレトロウイルスのプラスミドに頻繁に出現します。ただし、レトロウイルスは DNA をバイパスする代わりに、RT を使用してゲノムを宿主の DNA にエンコードします。
逆転写は、テロメア合成の形で、感染していない生殖系列真核細胞でも発生します。テロメアと呼ばれる染色体の末端では、酵素テロメラーゼが DNA の 1 本の鎖を長くします。テロメラーゼは、RNA の短い鎖をテンプレートとして使用してこれを行い、テロメアに結合した相補的 DNA の反復鎖を合成します。この延長により、DNA 複製中の DNA 染色体の短縮が防止されます。