コア コンセプト
この記事では、各 DNA 互変異性体 (B-DNA、A-DNA、Z-DNA) の構造と生化学的特性について学びます。
互変異性体とは?
DNAといえば、ねじれたはしごのような二重らせんのイメージがすぐに思い浮かびます。しかし、他の多くの有機分子と同様に、DNA は、特定の環境条件が与えられると、互変異性体と呼ばれるさまざまな形をとることができます。生化学者に B-DNA、A-DNA、Z-DNA として知られているこれらの DNA 異性体は、生物学的システムにとって重要な異なる化学的性質を持っています。

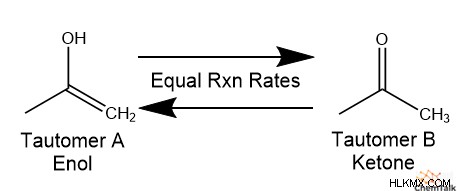
化学者は、「互変異性」を、2 つの分子構造が自発的に相互に変換される反応と定義しています。互変異性体と呼ばれるこれらの 2 つの分子は、構造異性体になる傾向があります。つまり、化学式は同じですが、原子間の結合が異なります。実際、多くの互変異性体は、水素原子の配置のみが異なります。また、互変異性体は動的平衡状態で存在する傾向があります。これは、互変異性体 A から互変異性体 B への変換率が、A と B の相対濃度が経時的に一定に保たれるように、A に変換する B の変換率と等しいことを意味します。
ケト-エノール互変異性は、有機化学および生化学における互変異性体ペアの最も一般的な例です。
他の互変異性体とは異なり、B-DNA、A-DNA、および Z-DNA は分子構造が異なるのではなく、2 つのヘリックス間の分子間相互作用が異なります。
B-DNA 互変異性体
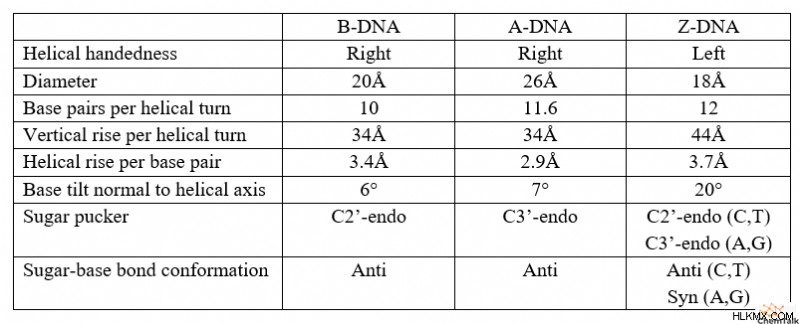
科学者もそうでない人も同様に、B-DNA は最も認識しやすい形の DNA として機能します。ロザリンド・フランクリンが X 線結晶学を使用して最初に画像化した B-DNA は、ワトソンとクリックの DNA モデルの基礎として機能しました。 B-DNA は、その利き手、塩基対の配置、およびシュガー パッカーによって、他の互変異性体とは異なります。
最初に、B-DNA 互変異性体が曲がり、「右巻き」の二重らせんを形成します。これは、時計回りに上向きにねじれる「左巻き」の二重らせんとは対照的に、鎖が反時計回りに上向きにねじれることを意味します。この区別を覚えるために、自分の右手が自分の前にあると想像してください。親指を上に向けて 、指を握りこぶし(または「親指」)に丸めます。指が反時計回りに曲がっていることがわかります .左手で同じことをすると、ファインダーが時計回りに曲がるのがわかります。 親指が上を向いているとき。
第二に、B-DNA の塩基対はヘリックス軸に対してほぼ垂直な平面を形成します。別の言い方をすれば、B-DNA の二重らせんを垂直に向けると、塩基対は DNA 鎖間にほぼ完全に水平な橋を形成します。
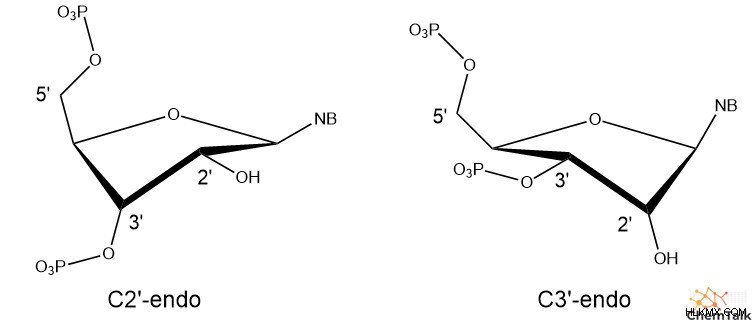
第三に、B-DNA は C2'-endo と呼ばれるシュガー パッカー構造を持っています。これは、DNA のデオキシリボース糖の 3D 立体構造を指します。具体的には、C2'-endo パッカーは、糖の 2' 炭素が 5' 炭素に結合したリン酸と同じ方向を向いていることを示します。この配向により、代替の C3'-endo と比較して、5' リン酸と 3' リン酸の間の距離が延長されます。
A-DNA 互変異性体
B-DNAほどの名声と認知度はありませんが、A-DNAはロザリンド・フランクリンの研究によって同時に発見されました。
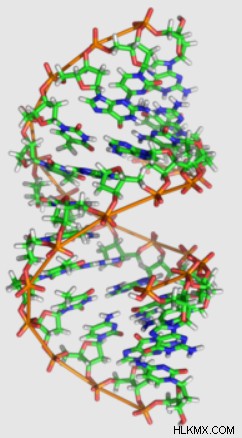
B-DNA と同様に、A-DNA も右巻きのらせん構造を持っています。ただし、A-DNA のデオキシリボースは C3'-endo のパッカーを示し、B-DNA よりも 5' および 3' リン酸を近づけます。これにより、A-DNA は B-DNA に比べて短くしゃがんだ外観になります。
さらに、らせん軸に対して垂直から20度の角度でA-DNAの塩基対が存在する。これらの角度のある塩基対は、B-DNA のように中心を横切って伸びるのではなく、らせんの外面に沿って鎖を接続します。これにより、二重らせんが「中空」になり、構造の内部にストローのような穴ができます。
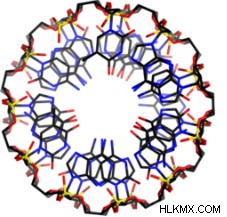
生化学者は、彼らが「脱水状態」と呼ぶ条件下で A-DNA が形成されることを発見しました。これは単純に、DNA と相互作用する水分子が比較的少ないことを意味します。近くに水がないと、「疎水性効果」が弱まります。これは、生体分子の極性領域が表面に引き寄せられ、非極性領域が内部で遮蔽される現象です。 B-DNA では、極性デオキシリボースとリン酸骨格がらせんの外側で水と相互作用します。逆に、窒素塩基の非極性領域は、B-DNA の安定性に重要な「スタッキング相互作用」を形成します。
A-DNA はらせんの外側に沿った塩基ペアリングを含むため、弱められた疎水性効果のみが A-DNA 互変異性体の形成を可能にします。
その発見以来、生化学者は、細菌が溶媒を除去するタンパク質を使用して A-DNA の形成を誘導できることを発見しました。おそらく、細菌がこれを行うのは、A-DNA互変異性体が極度の熱や乾燥の状態から遺伝子をよりよく保護するためです.
興味深いことに、二本鎖 RNA の構造は A-DNA の構造に似ています。
Z-DNA 互変異性体
ロザリンド・フランクリンの発見から数十年後、生化学者のアンドリュー・ワンとアレクサンダー・リッチは、特定の DNA 配列が Z-DNA と呼ばれる 3 番目の互変異性体を示すことを発見しました。
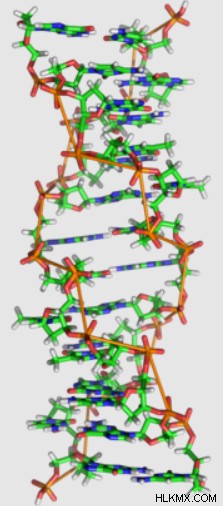
他の互変異性体とは異なり、Z-DNA は左巻きのらせん構造を持っています。 A-DNA および B-DNA と比較して、Z-DNA はより細長く、細いらせん構造を持っています。
興味深いことに、Z-DNA は、プリン (アデニンとグアニン) とピリミジン (チミンとシトシン) が交互になった DNA 配列間でのみ形成されることが観察されています。このような配列には、グアニンとシトシンが交互になった 2 本の鎖 (ポリ (GC) – ポリ (GC))、またはアデニンとシトシンの 1 本の鎖がグアニンとチミンの別の鎖と対になったもの (ポリ (AC) – ポリ (GT)) が含まれる可能性があります。いくつかの他のシーケンスの中で。
Z-DNA では、これらの配列は興味深い構造を想定しており、プリンには C3' エンド パッカーがあり、ピリミジンには C2' エンド パッカーがあります。さらに、糖とプリン塩基の間の結合は回転して syn コンフォメーションになり、塩基の環構造の大部分がデオキシリボースの上にぶら下がっています。 A-DNA、B-DNA、および Z-DNA ピリミジンでは、すべての糖と塩基の結合にアンチコンフォメーションがあり、塩基は糖の反対側を指しています。
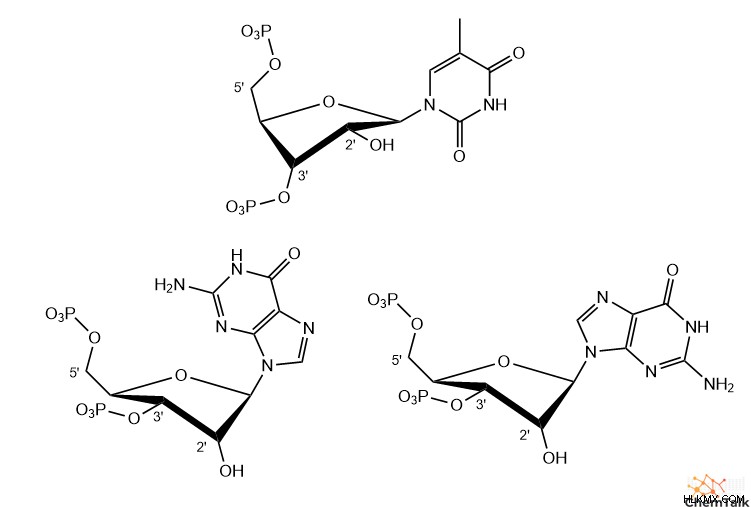
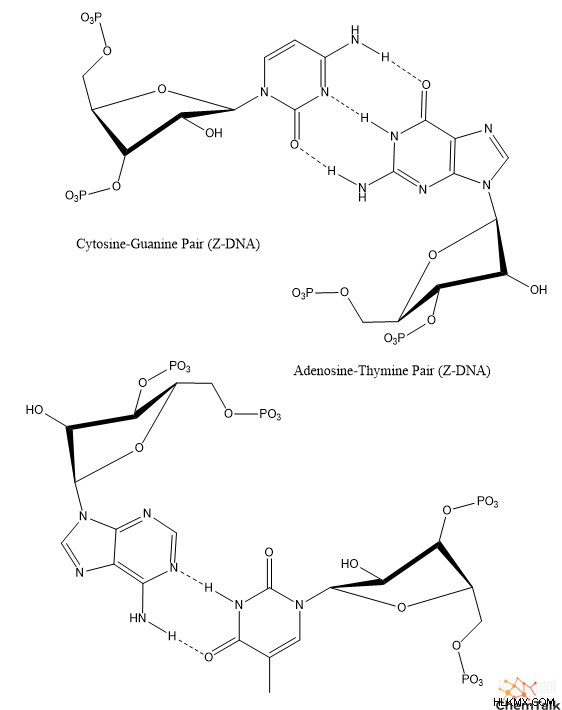
A-DNA と同様に、Z-DNA にも重要な生物学的目的があります。具体的には、酵素が転写のためにB-DNAを開いて巻き戻すと、Z-DNAが「負のスーパーコイル」を介して形成され、巻き戻しのストレスが軽減されます。実際、生化学者は、Z-アルファ ドメインなど、Z-DNA と特異的に相互作用する多くの異なる酵素と結合タンパク質を特定しています。
ただし、配列の仕様は別として、Z-DNA は高塩濃度下でのみ形成されることが観察されています。生化学者は、塩イオンが帯電したリン酸を安定化するのに役立つと信じています。これらのイオンがなければ、負に帯電したリン酸塩のイオン反発により、構造は安定性を失います。
DNA 互変異性体の概要表