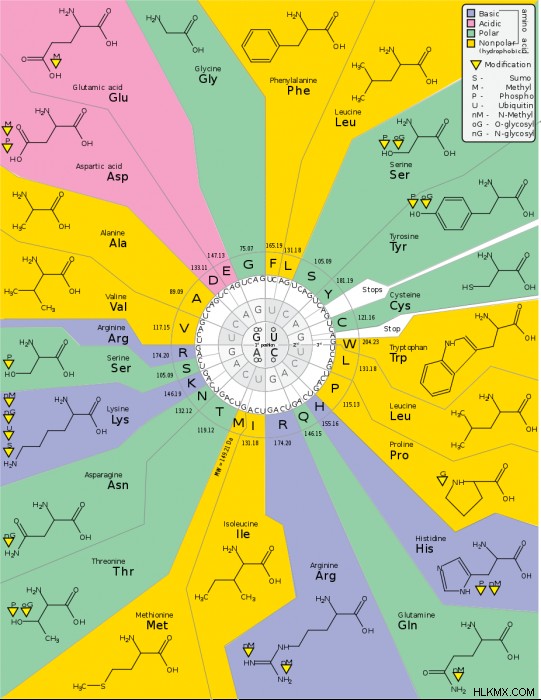
コドン チャートまたはテーブル DNAまたはRNAに対応するアミノ酸が使用されます。コドンチャートはポリペプチド鎖をまとめるのに役立ちますが、最初にコドンを知る必要があります.
ヌクレオチド 私たちのDNAを構成するものです。それは、私たちを遺伝的に形成するすべてのものを定義する言語です。コドンは、DNA の言語とタンパク質(アミノ酸)の言語をつなぐ翻訳者です。
ヌクレオチドは、糖、リン酸、および窒素塩基の 3 つの部分で構成される DNA のビルディング ブロックです。私たちの DNA には、アデニン(A)、グアニン(G)、シトシン(C)、チアミン (T)の 4 つの塩基があります。これらの 4 つの塩基は、さまざまな遺伝子、タンパク質、レギュレーター、および DNA が使用されるその他すべてをエンコードするために使用されます。
私たちのDNAは二重らせんなので、多くのヌクレオチドからなる鎖が2本あることになります。これらの 2 本の鎖は、それぞれの塩基が別の塩基と対を成すときに、これらのヌクレオチドを介して互いに結合します。塩基 A は T と対になり、塩基 G は C と対になり、逆もまた同様です。 DNA が RNA になると、チアミンがウラシル (U) に置き換わります。 RNA では、U は T の代わりに A とペアになります。
体内で生成される多くの異なる RNA の中で、タンパク質合成の目的で生成されるものの中には、タンパク質の特定の側面を表す 3 桁のコード行を含むものがあります。これらの 3 桁のコードはコドンと呼ばれ、タンパク質が適切に合成されることを保証する重要な部分です。
コドンとタンパク質合成
コドンは、4 つの RNA 塩基の任意の組み合わせで構成される 3 桁の数字です。 DNA コドンは、U の代わりに T があることを除いて同じです。表を見ると、合計 64 のコドンがあり、それぞれ特定のアミノ酸またはタンパク質合成のための機能を表しています。
タンパク質合成では、メッセンジャー RNA (mRNA) がタンパク質と RNA からなる構造体であるリボソームに到達し、タンパク質を作成するプロセスを開始します。 mRNA には、リボソームが「読み取り」、タンパク質を作成する編集されていないタンパク質の配列が含まれています。
一般に、AUG、メチオニン、コドンの位置は、タンパク質合成の開始を示します。リボソームが開始コドンを見つけると、トランスファー RNA (tRNA) を取り込みます。この tRNA には、アンチコドンとそれに結合したアミノ酸が含まれています。アンチコドンは、単にmRNAのコドンに相補的な配列です。したがって、AUG は UAC によって補完されます。
タンパク質合成が機能するには、mRNA のコドンが tRNA のアンチコドンと一致する必要があります。一致すると、リボソームは一度に 2 つの tRNA を保持できるため、次の tRNA を取り込み、次のコドンと一致させます。一致する場合、この新しい tRNA のアミノ酸は既存のアミノ酸に結合され、リボソームは次のコドンにシフトします。
このプロセスは、リボソームが mRNA 上のストップ コドン (UAA、UAG、または UGA のいずれか) に遭遇するまで続きます。これら 3 つのいずれかがタンパク質合成を停止します。この時点で、合成されたタンパク質が解放され、処理を完了するために送信されます。
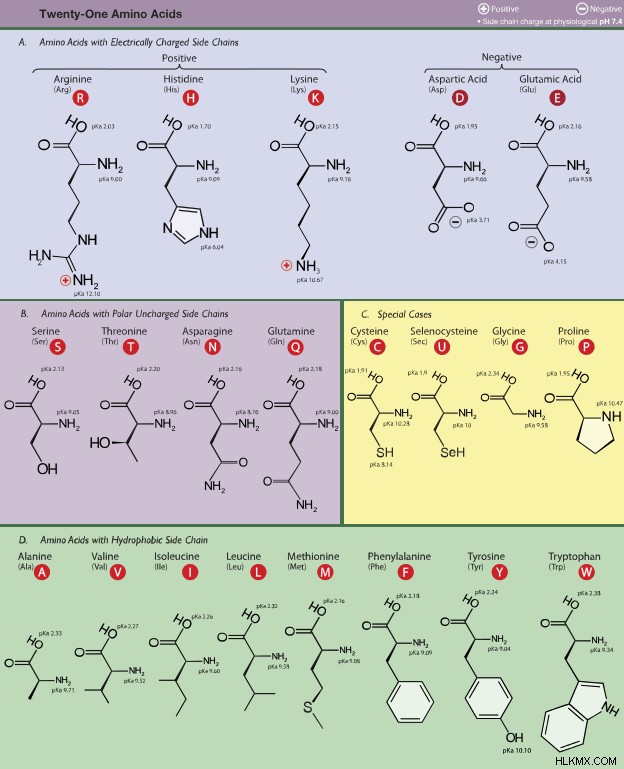
停止コドンは 3 つあるため、残りのコドンのうち 61 個がアミノ酸をコードしていることになります。人間のアミノ酸は 20 個しかないため、コドン内には多くの重複があります。たとえば、UUA と UUG は両方ともロイシンをエンコードします。 CCU、CCC、および CCA はすべてプロリンをエンコードします。
冗長性と変異
コドンは、タンパク質合成を混乱させる可能性のある突然変異や問題に対する予防策として、非常に多数存在します.私たちの体の突然変異は、さまざまながんや遺伝性疾患につながる可能性があるため、問題になる可能性があります.いくつかの突然変異は、私たちを多くの病気や非遺伝性疾患の危険にさらすことさえあります.
リスクは常に存在しますが、コドンの冗長性により、これらの変異を最小限に抑えることができます。
同義の置換
同義置換は、特定のタンパク質のアミノ酸を表す DNA 配列の突然変異であるため、サイレント突然変異と呼ばれることが多く、アミノ酸を別のものに変更することはありません。たとえば、DNA では CTT はロイシンになります。同義の突然変異はそれを CTC に変更しますが、これはまだロイシンをコードしているので問題はありません.
現在、これらの同義変異は、複数のコドンを持つアミノ酸に対しては一般に無害ですが、特定のアミノ酸に対して特定のコドン配列を持つことを好む種があることが示されています.
コドンの冗長性により、これらの変異は無視でき、元のコドン配列と同じアミノ酸をコードしている限り害はありません。ただし、すべてのコドンが冗長な形式を表すわけではありません。 ATG のみで表されるメチオニンは、それをコードするコドンが他にないため、「サイレント」変異によって容易に別のものに変異する可能性があります。そのため、ATG を ATA に変更すると、メチオニンがまったく異なるアミノ酸であるイソロイシンに変更されます。
非同義置換
メチオニンへの変化の可能性と同様に、すべての突然変異がサイレントで無害というわけではありません.害を及ぼすものは、一般に非同義置換 (または突然変異) と呼ばれます。これらは、タンパク質のアミノ酸を変更するために発生する可能性のあるさまざまな問題をカバーしています.
たとえば、CTT (ロイシン) が CCT (プロリン) に変更されると、タンパク質に根本的で顕著な変化が生じます。これはしばしばミスセンス変異と呼ばれます。
置換の別の形態は、アミノ酸コドンのストップコドンへの変化をもたらすナンセンス突然変異であり、タンパク質合成プロセスを時期尚早に終了させる.
これらは、コドン配列を変更し、それらが合成するタンパク質に潜在的に有害な影響を与える可能性のある変異の一部です.他のものは、タンパク質の一連のコドンから特定のヌクレオチドを削除する有害な突然変異である可能性があります.リボソームは特定の時点で mRNA の「読み取り」を開始するため、削除によって開始位置が変更され、変更されたタンパク質産物が作成されます。同様に、一連のコドンにヌクレオチドを挿入する挿入変異は、リボソームが mRNA の「読み取り」を開始する場所を変更します。