ミトコンドリアは、動物、植物、菌類の細胞内にある小さな体で、細菌や古細菌の細胞が関与する内部共生プロセスに由来します。その小さなサイズにもかかわらず、ミトコンドリアはエネルギー生産、ストレス応答、およびその他の重要な生化学的経路に不可欠です。これらのプロセスは忙しい工場のように見えますが、他の工場と同じように、生産量が多いほど無駄が多くなります.
通常の活動を行うために、ミトコンドリアはほとんどの場合、独自のタンパク質を生成しません。それらのほとんどはオルガネラの外側で合成され、通常は N 末端ターゲティング ペプチドに存在する選別シグナルの助けを借りて内部にインポートする必要があります。前駆体タンパク質がインポートされた後、ターゲティング ペプチドはプロセシング ペプチダーゼによって切断され、副産物としてミトコンドリア マトリックスに放出されます。他の廃棄物と同様に、フリー ターゲティング ペプチドは適切に処理しないと危険です。
ヒトでは、遊離ペプチドの蓄積が多くの疾患に関連しています。悪名高い例は、アルツハイマー病を引き起こすアミロイド ベータ ペプチドの蓄積です。植物では、ペプチドの蓄積はオルガネラの安定性の破壊と関連していました。これは、ペプチドが強く相互作用して脂質膜を破壊する可能性があるためです。遊離ペプチドは、プロセッシング ペプチダーゼに結合し、時期尚早のストレス反応を引き起こす可能性があるため、タンパク質のインポート プロセスを妨害する可能性もあります。
ミトコンドリアは、この廃棄物の問題を管理する方法を進化させてきました。非常に詳細に解明されたメカニズムの 1 つは、植物で最初に完全に特徴付けられた処理経路です。
プロテアソームによるユビキチン化タンパク質またはペプチドの直接的な分解であるサイトゾルでのプロテアソーム分解とは対照的に、処理経路には複数の異なるペプチダーゼによる段階的な分解が必要です。長い遊離標的ペプチドは、プレタンパク質ペプチダーゼ (PreP) によってオリゴペプチドに断片化されます。オリゴペプチドは、オルガネラのオリゴペプチダーゼ (OOP) によってさらに分解され、短いペプチドになります。最後に、アミノペプチダーゼによって短いペプチドから単一のアミノ酸が放出されます。
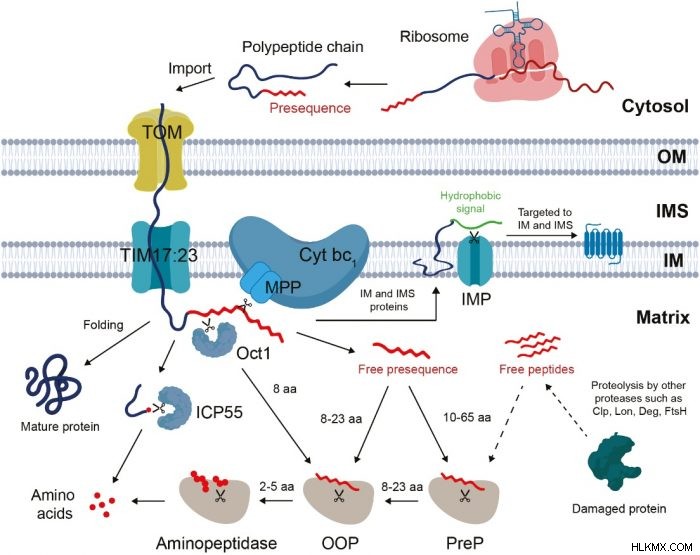
興味深いことに、このペプチド分解メカニズムは、ミトコンドリアと植物の別の内部共生オルガネラである葉緑体との間で共有されています。 PreP、OOP、アミノペプチダーゼなど、プロセッシング経路に関与するペプチダーゼのほとんどは、二重標的化されています。つまり、これらのペプチダーゼは、ミトコンドリアと葉緑体の両方を標的とし、局在しています。これらのペプチダーゼの不活性化は、オルガネラの安定性と活性を変化させ、植物の成長、発育、ストレス応答に全体的に影響を与えます.
植物には何百もの異なるプロテアーゼとペプチダーゼがあるため、植物の成長と環境条件への応答に不可欠なプロセスに他のタンパク質が関与している可能性があります.