騒ぎを引き起こす方法は次のとおりです。多くのナチュラリストに世界を単純化するよう依頼します。私たちは通常、動物、植物、微生物、地球、風、火の間の複雑な相互作用のネットワークの観点から考えています。これは、ダーウィンが「絡み合った土手」と呼んだものです。銀行の複雑さを一般化して単純化することは、不誠実に思えるかもしれません。
したがって、ウィスコンシン大学の理論生態学者であるトニー・アイブスが、2013 年のアメリカ生態学会の会議で、同僚が一般的な法則を追求すべきかどうかについて投票するよう求めて、同僚をせき立てたとき、おそらく驚くべきことではありませんでした。 -会議室の 3 分の 1 が反対票を投じました。
懐疑論があるにもかかわらず、単純化によって可能になった種類の一般法則には、驚くべき予測力があります。大きすぎて完全にサンプリングできない生態系にどれだけの種が存在するか、または生息地の破壊後にどれだけの種が失われるかを計算できるようになる可能性があります。
物理学を学んだ後に生物学を始めたせいか、私はこれらの一般的な法則に美を見出している生態学者です。物理学では、気体の分子と別の分子との違いは最も気にしないものです。個人的に好きな電子を持っている人はいません。圧力、体積、および温度に関連する理想気体の法則は、酸素と窒素にも同様に当てはまります。液体と気体の間の相転移は、特定の金属の磁化と同じように振る舞います。
生態系が理想気体のように美しく完全であってはならないのはなぜでしょうか? また、生態学者が物理学者ほどの予測力を持てないのはなぜでしょうか?これらの質問への答えは、「そうです」と「できる」かもしれません。ただし、特定の視点から見た場合のみです。
1980 年代、ニューメキシコ大学のジム・ブラウンとブリガム・ヤング大学のブライアン・マウラーという 2 人の生態学者がマクロエコロジーという用語を作り出しました。マクロエコロジーは、自然界の創発的パターンを探求する研究者に名前と知的な家を与えました。多くの生態学的研究が小規模であることに不満を感じていたマクロエコロジストは、自然を時間と空間で広く説明できるパターンと理論を探していました。
ブラウンとマウラーは、多くの生態学的現象の規則性に大きな影響を受けていました。種面積曲線と呼ばれるこれらの 1 つは、19 世紀に発見され、1921 年に正式化されました。この曲線は、自然主義者が敷設された区画で見つかった (植物、昆虫、哺乳類などの) 種の数を数えたときに現れました。裏庭、サバンナ、森で。彼らは、予想通り、プロットの面積とともに種の数が増加することを発見しました。しかし、区画サイズが大きくなるにつれて、種数の増加率は頭打ちになり始めました。さらに驚くべきことに、種や生息地に関係なく、同じ基本的な種-面積曲線が見つかりました。数学的に言えば、曲線は累乗則に従い、種数の変化は面積の平方根の平方根に比例して増加しました。
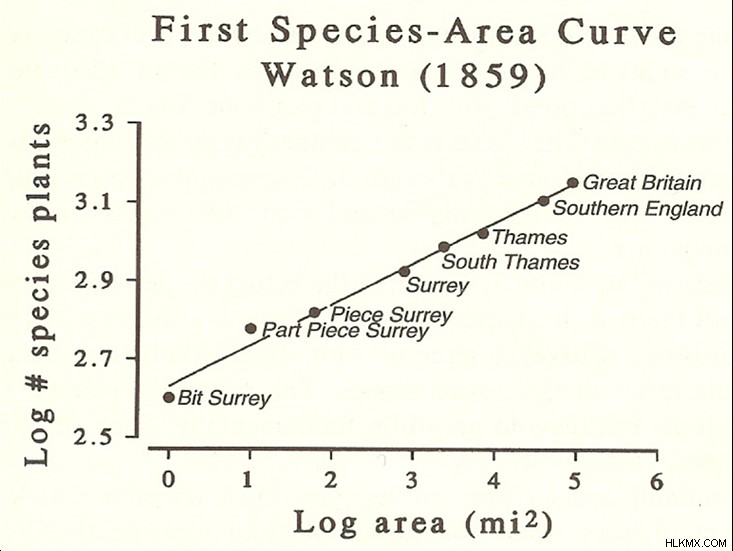
べき法則は科学では一般的であり、物理学における普遍性の決定的な特徴です。彼らは、気温の上昇に伴う磁石の強さ、サイズに対する地震の頻度、人口の関数としての都市の生産性について説明しています。多くの生態学者にとって、種面積曲線は神経質になります。これは、十分に大きなスケールでは、生態系の特定の詳細 (生態学者の心にとって非常に近く、大切な「絡み合った土手」) は単に問題ではないことを示唆しています。特異性は洗い流され、生態系は他の科学のさまざまなシステムに驚くほど似ているように見え始めます。
種面積曲線の普遍性は、生態学者がそれを理解するために構築したモデルで非常に明確になりました。 2001 年、当時プリンストン大学の生態学者であったスティーブ ハッベル (Steve Hubbell) は、食物連鎖の同じ部分にいる個体間の選択的差異 (進化上の利点または不利益を与える差異) は無関係であるという根本的な仮定に基づいたモデルを開発しました。つまり、個々の結果は一種のエコロジカル ルーレットです。いくつかの種は幸運に恵まれ、最終的に宇宙全体に広く豊富に分布するようになりますが、他の種は比較的まれになります.分析方程式とコンピューター シミュレーションを組み合わせることで、生物多様性の統一中立理論と呼ばれる彼のモデルは、驚くほど現実的に見える種面積曲線を予測しました。その成功は、植物、動物、生物がほぼ同一の統計的プレースホルダーに置き換えられた、実際の生態系のこの残酷に単純化されたバージョンに基づいています。
物理学の訓練を受けた別の生態学者であるジョン・ハートは、中立的な理論が想定するよりもさらに少ない生態学的メカニズムで種面積曲線を理解できるかどうか疑問に思いました。 Harte は、熱力学と情報理論から得られたアイデアに基づいて、生態学の最大エントロピー理論を開発しました。エントロピーはシステムの無秩序の尺度であり、熱力学で使用され、固定体積内の同一のガス分子の最も可能性の高い配置を計算します。通常、より多くの無秩序が勝ちます。特定の制約の下で種の空間分布を操作することにより、Harte は最大エントロピー理論を使用して、インドの西ガーツ山脈全体の樹木種の数を予測しました。 Ecology Lettersに掲載された彼らの見積もり 、カウントされた約900種類の木のかなりの10%以内に収まりました。 Harte は、個々の木の詳細や、その繁殖や種子の分散については考慮していませんでした。彼の研究は、情報理論の領域からの原則によって純粋に推進されていました。
種が互いにどのように競合し、どのように異なるかの詳細を無視することにより、最大エントロピーと中立理論は、生態系の乱雑で複雑なもつれを、理想的な気体の理想化された完全性に変換します。そうすることで、生態学者は物理学のような予測と説明の能力を得ることができます。しかし、どちらのモデルも物議を醸しています。これらの詳細は、まさに生態学者が一生をかけて調査できるものであり、それらが問題ではないことを示唆する 2 つの理論があります。
これらの空間予測が正しくても、理由が間違っていたとしたら?おそらく、一般的な生態学的モデルには実際の生態学的詳細とメカニズムを含める必要があり、これらの詳細を無視しても成功するモデルはまぐれであり、種面積曲線のような空間パターンだけに制限されています.発見する 1 つの方法は、これらのモデルを別の次元に拡張することでした。私は時間を選びました。
生態学者は、50年以内に何種類の樹木が死ぬかを推定するために何をしますか?何千年も前にいくつの血統があったでしょうか?種面積曲線をモデル化するものと同様の理論は、空間ではなく時間だけに焦点を当てて、便利になるでしょう。そこで私は、以前私と協力していた同僚であるトム・シャープトンとスティーブ・ケンベルを採用して、1 つを構築しました。
細菌は、信じられないほど豊富に存在することと、DNA 配列データによって進化の歴史を知ることができるという理由から、当然のように思われました。私たちの計画は、種面積曲線で行われたのと同じように、種の数を数えることでした。ただし、面積の代わりに、時間の尺度を使用します。
私たちは、バクテリアの DNA 配列を比較し、生命の木を描くことで、その尺度を構築しました。木の各枝は、新しいバクテリアの系統を表していました。これは、過去の生命のある種の多様化です。ツリー上の種間の平均進化距離 (または枝の長さ) は、時間をかけてそれらの関連性を定量化しました。私たちがサンプリングした微生物は、人間の鼻腔、人間の糞便、植物の葉の表面、南極海、イギリス海峡の水など、約 25 の異なる生息地から採取されました。
平均進化距離を種数に対してプロットしたところ、生態学のさらに別の次元に潜むべき法則が見つかりました。距離は、最初は急速に増加し、その後、種面積曲線と同じように減速し始めました。この動作の理由は現時点では明らかではありません。 1 つの可能性は、空間的および時間的なスケーリング動作の両方が「バースト性」の影響を受けることです。バースト性では、スタシスの期間が急速な多様化の期間によって中断されます。私たちのバクテリア ツリーでは、これらのバースト展開がべき法則によっても記述されるフラクタル分布を持ち、時間と空間の両方で種の放射を示す可能性があることを発見しました。
進化の距離と多様化について私たちが目にするべき法則は、最大規模での生態学の単純で機械的で、比較的詳細のない見方を再び指し示しています。それらは、空間パターンに対して提案されているものほど単純ではありません。彼らは、マクロ生態学的パターンを説明するために真の生態学的および進化的メカニズムを必要とする方向に、スペクトルを少なくとも一歩後退させます.
別の生態学者が私に尋ねたことがあります。スケーリングから何を学ぶことができますか?それは公正な質問です。種面積曲線を使用したことがない生態学者はたくさんいます。私にとっては、知的なスリルと挑戦、そして異なるシステム間で予想外の類似点を見つけたときの驚きの要素があります。また、スケーリングによって可能になる予測の力もあります。マクロ生態学を理解するのに本当に役立つ理論を開発できれば、今日のアマゾンの多様性の喪失が100年後に熱帯地方をどのように変化させるかをより正確に見積もることができるかもしれません.または、人間の腸内の細菌の相互作用がその未来をどのように変えるか.
さらに何かがあります。生態学では、昆虫から鳥、草からセコイアまで、常に巨大な異質性が存在します。集約されたパターンだけでなく、さまざまな生物や種の違いを気にすることができます。したがって、生態学における普遍的で単純なスケーリング パターンについて学ぶとき、私たちはまったく新しい課題に直面しています:集約されたパターンでのスケーリングから学んだ一般性を、自然史やさまざまな生物やシステムの細部とどのように統合するのでしょうか?
Ecological Society の会議での Ives の質問に対する有用な補足があります。エコロジーが一般的な法則を追求すべきかどうかだけでなく、それらの法則を追求する私たちと、それらを生み出すもつれに焦点を当てる私たちとがどのように協力できるかということです.そのつながりを理解するとき、学ぶべきことはたくさんあります。
James O’Dwyer は、イリノイ大学アーバナ シャンペーン校植物生物学科の理論生態学者です。
参考文献
1. Fox, J. ESA Monday review:Tony Ives rocks. ダイナミック エコロジー dynamicecology.wordpress.com (2013).
2. Harte, J.、Smith, A.B.、および Storch, D. 生物多様性は、普遍的な種面積曲線を使用してプロットからバイオームにスケーリングします。 エコロジーレター 12 、789-797 (2009)。
3. O’Dwyer、J.P.、Kembel、S.W.、および Sharpton、T.J.進化の歴史のバックボーンは、微生物の生物多様性理論をテストします。 全米科学アカデミーの議事録 112 、8356-8361 (2015).
リード画像は Edhv, www.edhv.nl (オランダ、アイントホーフェン) の厚意によるものです。