中心的なコンセプト
この記事では、ペニシリンのような β-ラクタム系抗生物質が生化学レベルでどのように作用するか、さまざまな形態のペニシリンの特徴、そしてペニシリンが微生物学の分野をどのように再形成したかについて学びます。
ペニシリンの発見
歴史上最も影響力のある医学的発見の中には、偶然に生まれたものもあります。 20 世紀初頭、細菌性疾患が蔓延し、アレクサンダー フレミングのような生物医学者は、この種の感染症を理解し、これと戦うことに研究を捧げました。
フレミングのブドウ球菌の研究 、細菌の属であるため、ペトリ皿上で培養する必要がありました。 1928 年のある例では、 彼はブドウ球菌から離れました。 旅行中に皿の上で成長するために。休暇の後、彼は研究室で珍しい観察に戻りました。 ペニシリウム すでに細菌が増殖していた 1 つの皿が真菌 (カビ) によって部分的に汚染されていました。その皿では、真菌を取り囲んでいた細菌は死んでいましたが、同じ皿上の真菌から遠く離れた他の細菌のコロニーは繁栄していました。
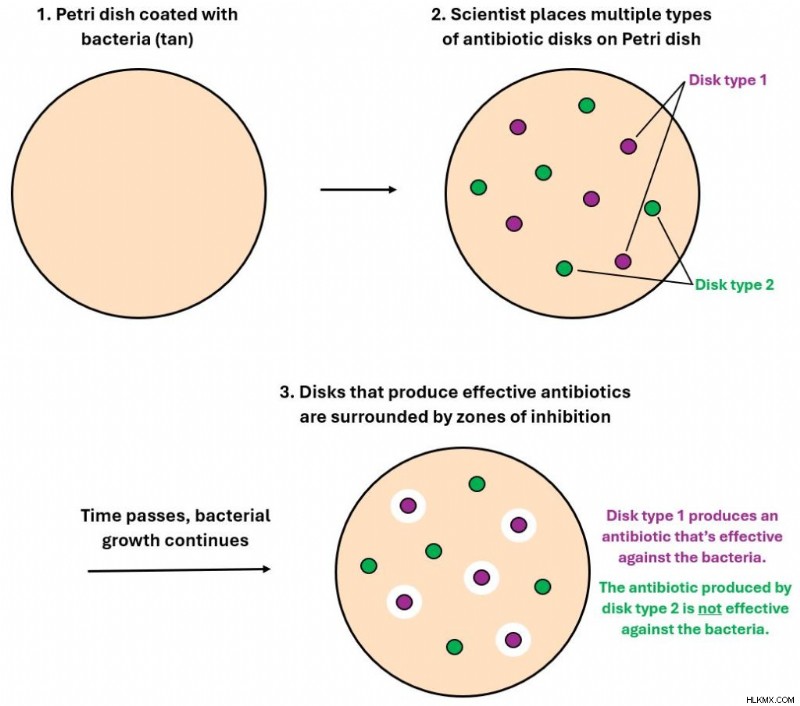
皿に抗生物質が存在しないと、細菌が自由に増殖してしまいます。しかし、抗生物質のディスクが入った皿で細菌が増殖すると、その影響は明らかです。抗生物質がこの種の細菌に対して効果がある場合、その抗生物質ディスクが周囲の細菌を殺し、 その結果抑制ゾーンが形成されます。 。このゾーンは、細菌が根絶されているため、細菌の増殖が存在しないペトリ皿上の領域です。抗生物質が存在していても、このタイプの細菌に対して効果がない場合、細菌は抑制ゾーンなしで抗生物質ディスクの隣で増殖し続ける可能性があります。
この観察に基づいて、フレミングはペトリ皿に使用された培養液には細菌を殺す作用(殺菌作用がある)があると結論付けました。 ) の性質を彼はペニシリンと名付けました。 。彼はまた、この菌が自然にペニシリンを生成し、この化合物をブロス中に分泌していることも突き止めた。さらなる研究を通じて、彼は、ブロスが一部の細菌種のみを殺すことができ、他の細菌種は影響を受けないことを示しました。ペニシリンを精製するための献身的な努力にもかかわらず、彼は成功せず、代わりに、後に彼の研究を基礎とする他の研究者と菌を共有しました。
β-ラクタム系抗生物質はどのように作用しますか?
ペニシリンが抗生物質であることがわかりました。 、ある種の病原性細菌を標的にして殺す薬の一種。フレミングの研究はペニシリンについて私たちが知っていることの基礎となっていますが、エドワード・エイブラハム、エルンスト・チェイン、ドロシー・ホジキンによるその後の研究により、ペニシリンの抗生物質機能の基礎となる構造成分が決定されました。ペニシリンの殺菌能力の背後にあるメカニズムを調べてみましょう。
ペニシリンの生化学
ペニシリンはβ- ラクタムに属します。 抗生物質の一種で、分子構造にβ-ラクタム環があることを意味します。この環は、窒素原子がカルボニル基の β 炭素原子に結合している環状アミドを特徴としています。
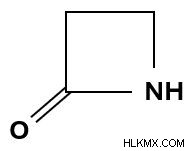 β-ラクタム環。
β-ラクタム環。 ペニシリンでは、この窒素原子は、β-ラクタム環に隣接するチアゾリジン環の一部でもあります。この結合構造はペナムリングとして知られています。 。ペニシリンには、そのさまざまな形態のすべてに常にペナム環が含まれています。ただし、ペニシリンの独特の形態を区別するのは、側鎖内の異なる R 基です。
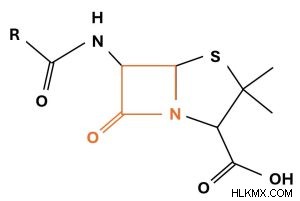 β-ラクタム環がオレンジ色の枠で囲まれたペニシリン分子。 R 基はペニシリンの形態によって異なります。
β-ラクタム環がオレンジ色の枠で囲まれたペニシリン分子。 R 基はペニシリンの形態によって異なります。 ペプチドグリカン
2 種類の細菌 (マイコプラズマを除く) およびL型細菌)には細胞壁があります。細胞壁を有する細菌では、 細胞壁はペプチドグリカンで作られた外膜によって強化されています。 。各ペプチドグリカン モノマーは 2 つの糖で構成されます:N - アセチルムラミン酸 (ナム )、これもテトラペプチドに結合しており、 N ・ アセチルグルコサミン (ナグ )。テトラペプチド内の 4 つのペプチドは、細菌種によって異なります。個々のモノマーのテトラペプチド鎖が架橋して、細菌細胞の外側の周りにメッシュ状の結晶格子を形成します。
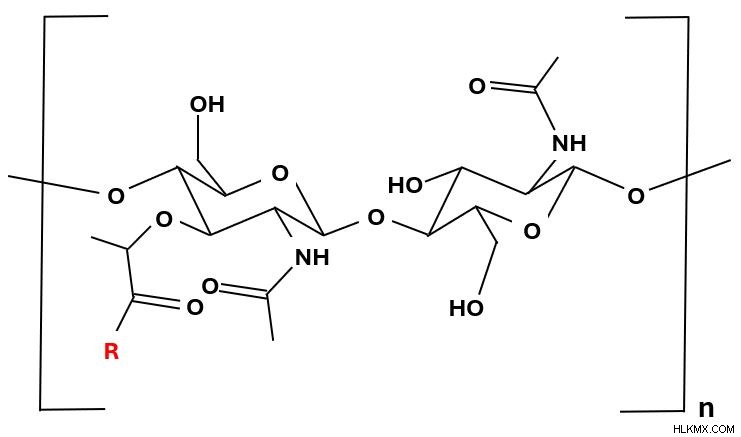 ペプチドグリカン モノマー。 R 基 (赤色) はテトラペプチドを表します。
ペプチドグリカン モノマー。 R 基 (赤色) はテトラペプチドを表します。 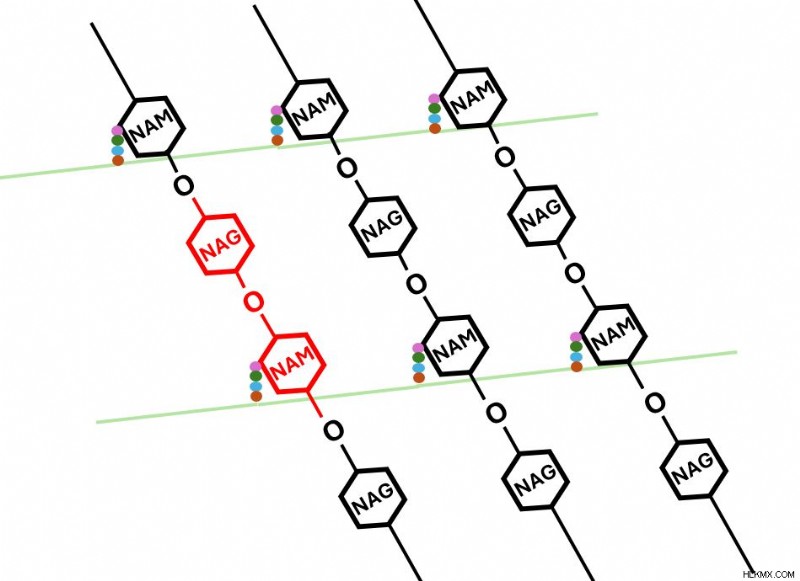 モノマーが赤色で強調表示されているペプチドグリカン ポリマー。各モノマーの NAM に結合しているテトラペプチド (多色丸) に注目してください。平行モノマーのテトラペプチド鎖が架橋 (緑色) し、格子を形成しています。
モノマーが赤色で強調表示されているペプチドグリカン ポリマー。各モノマーの NAM に結合しているテトラペプチド (多色丸) に注目してください。平行モノマーのテトラペプチド鎖が架橋 (緑色) し、格子を形成しています。 ペプチドグリカンは細菌細胞を三次元の足場のような構造で包み込み、細胞の構造を維持します。 β-ラクタム系抗生物質は、厚いペプチドグリカン層よりも薄いペプチドグリカン層のほうが容易に浸透できるため、 薄いペプチドグリカン層を持つ細菌細胞は死滅しやすいのです。 ペニシリンがこの層に浸透すると、細菌細胞の膜を内側から破壊します。しかし、これはどのようにして実現されるのでしょうか?
細胞壁のある細胞には膨圧があります。 (静水圧)は、細胞膜を細胞壁に押し付けて細胞の形状を維持する力です。この力は、細胞の内側と外側の間の浸透圧勾配から生じます。低張状態で観察されるように、浸透圧勾配の不均衡により細胞内に過剰な水が流入すると、 細胞が溶解する可能性があります。 (破裂して)そして死ぬ。溶解を防ぐには、膨圧による外向きの力に対抗するために、細胞壁を内向きに押す別の力が必要です。この内向きの力はペプチドグリカン層によって提供されます。ペプチドグリカンは細胞壁を強化するため、厚いペプチドグリカン層を持つ細菌細胞は溶解の脅威に対してより回復力があります。
ペニシリンはどのようにしてペプチドグリカン層を標的にするのですか?
DD-トランスペプチダーゼ は、テトラペプチドを架橋してペプチドグリカン層を合成する重要な酵素です。 β-ラクタム系抗生物質は、そのβ-ラクタム環がDD-トランスペプチダーゼの活性部位に結合し、酵素を阻害してペプチドグリカンの形成を防ぐため、病原性細菌に対して効果的です。ペプチドグリカンがないと、膨圧が細胞ウェルにかかる内向きの力に打ち勝つため、細菌細胞の外膜が弱くなります。最終的に、この膜は非常に弱くなり、細胞の内側と外側の間の濃度勾配を維持できなくなります。過剰な水が細菌細胞に入り、細胞死を引き起こします。
したがって、ペニシリンのようなβ-ラクタム系抗生物質は、細胞壁を欠く細菌によって引き起こされる感染症の治療には効果がありません。これらの細菌は浸透圧の不均衡の結果として溶解しないため、 溶解しないクラスの抗生物質のみが使用されます。 標的細胞壁の合成により細胞が死滅する可能性があります。医療専門家は、これらの複雑な細菌をうまく中和するために、さまざまな抗生物質を組み合わせて使用する必要がある場合があります。
ペニシリンはどのように合成されますか?
ペニシリンには多くの種類があります。以下に示すアモキシシリンやアンピシリンなどは、すでによく知られているかもしれません。 β-ラクタム環はあらゆる形態の中心ですが、合成プロセスのわずかな違いにより R 基が変化し、その結果、さまざまな形態のペニシリンが生成されます。たとえば、R 基がベンジル基の場合、 これをペニシリン Gと呼びます。 ですが、酸素原子が 1 つ追加されているだけで、 それはペニシリン V として区別されます。 代わりに。ただし、β-ラクタム環が実際に DD-トランスペプチダーゼを阻害する基であることを思い出してください。ペニシリン分子内の他の構造基(可変 R 基など)は、酵素に積極的に結合しません。
では、なぜ独自の形態のペニシリンを作成する必要があるのでしょうか?ペニシリンは万能ではありません。それぞれの形態は、異なる細菌との闘いに成功します。
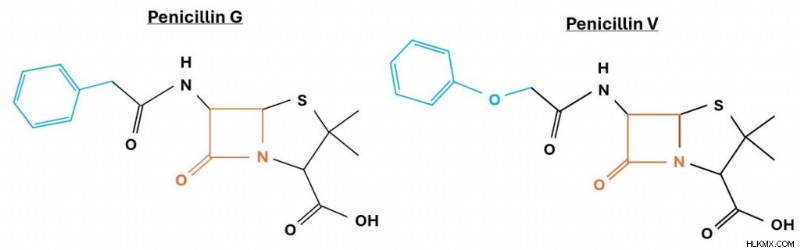 ペニシリン G 分子とペニシリン V 分子の比較。
ペニシリン G 分子とペニシリン V 分子の比較。 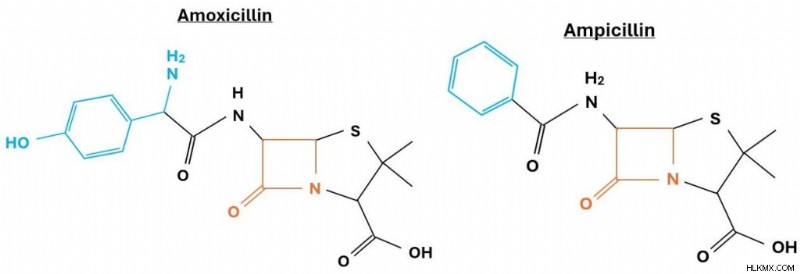 アモキシシリン分子とアンピシリン分子の比較。
アモキシシリン分子とアンピシリン分子の比較。 一般に、 ペニシリンはペニシリウムを発酵させることによって大規模に製造されます。 ペニシリン G や V などの β-ラクタム系抗生物質を自然に生成する真菌。カビが十分な量のペニシリンを生成すると、科学者はカビの混合物からペニシリン生成物を分離します。あるいは、特定の形態のペニシリン (アモキシシリンやアンピシリンなど) は、これらの天然分子の修飾バージョンです。製品が製薬業界の厳格な品質基準を満たしたら、医薬品として使用できるようになります。
ペニシリン V の生合成
ペニシリウムがどのように発生するかを分析しましょう ペニシリン V を自然に合成します。まず、L-α-アミノアジピン酸 (α-AAA)、L-システイン、D-バリンの 3 つのアミノ酸から始めます。酵素 ACV シンテターゼの助けにより、これらのアミノ酸は縮合反応を起こし、δ-(L-α-アミノアジピル)-L-システイン-D-バリン (ACV トリペプチド) と呼ばれるトリペプチドを形成します。
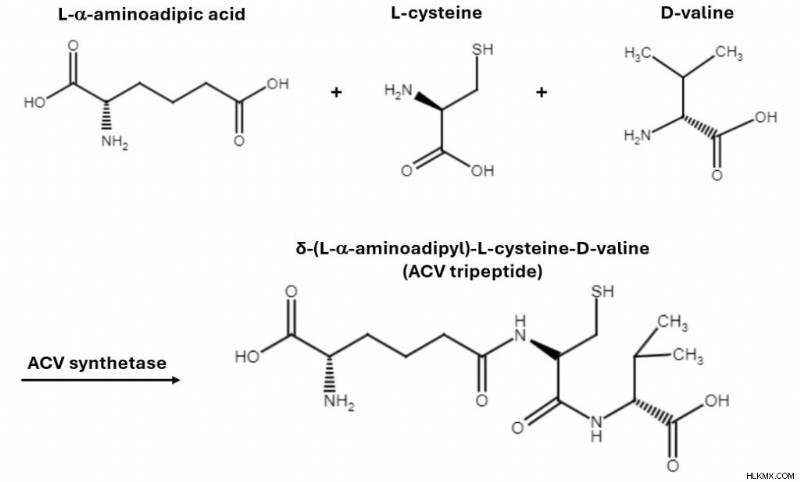 3 つのアミノ酸が凝縮して、ACV トリペプチドとして知られる単一の分子になります。
3 つのアミノ酸が凝縮して、ACV トリペプチドとして知られる単一の分子になります。 次に、酸化を通じて、酵素イソペニシリン N シンターゼ (IPN シンターゼ) が ACV トリペプチドをイソペニシリン N に変換します。このステップにより、β-ラクタム環が閉じられます。これはペニシリンの殺菌特性に重要であることがわかっています。
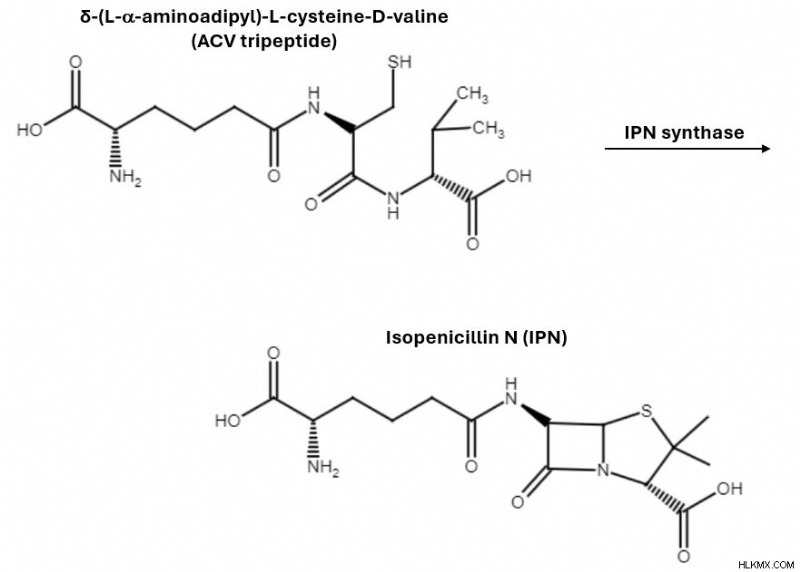 ACV トリペプチド中間体から始まり、IPN シンターゼは酸化反応を通じてイソペニシリン N の生成を触媒します。
ACV トリペプチド中間体から始まり、IPN シンターゼは酸化反応を通じてイソペニシリン N の生成を触媒します。 最後のステップでは、フェノキシアセチル CoA とアシルトランスフェラーゼを合成に導入します。これらの成分はアミド交換反応を実行し、IPN の α-AAA 部分が分子の残りの部分から分離されます。 CoA は IPN のカルボニル基を活性化し、CoA のフェノキシアセチル基を置換基として受け入れる準備をします。最後に、フェノキシアセチル基を IPN に寄付すると、CoA は反応から離脱し、標的分子であるペニシリン V を残します。
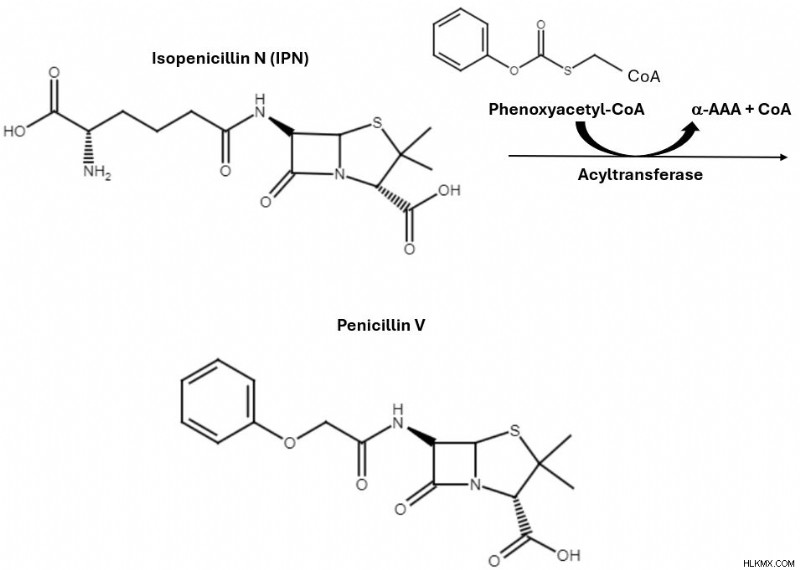 アミド交換反応によりペニシリン V の生成が促進され、その過程でフェノキシアセチル基が獲得され、α-AAA と CoA が失われます。
アミド交換反応によりペニシリン V の生成が促進され、その過程でフェノキシアセチル基が獲得され、α-AAA と CoA が失われます。 この合成によりペニシリン V が生成されましたが、化学者はこのメカニズムをさらに変更して、研究室で他の形態のペニシリンを生成することができます。 ペニシリウムの場合 自然界ではペニシリン G が生成され、これと同じ経路をたどりますが、代わりにフェニルアセチル CoA が使用されます (ここで見られるフェノキシアセチル CoA 反応物よりも酸素原子が 1 個少ない)。フェニルアセチル CoA の使用は、ペニシリン V の代わりにペニシリン G を作成するために必要な唯一の変更です。理論的には、別の形式のペニシリンを作成するために、任意の R 基を CoA のフェニルアセチル基またはフェノキシアセチル基と置き換えることができます。
ペニシリン V の合成:半合成ルート
半合成 は、出発物質が自然界に存在する化合物である合成アプローチです。これは合成化学者にとって近道として機能します。出発物質を自分で生成するのではなく、出発物質が自然に発生した場所 (細胞培養など) から出発物質を単離するだけで済みます。
ペニシリン V を半合成的に製造するには、ペニシリン G を出発物質として使用します。これはペニシリウムの一種であることを忘れないでください。 自然に生成します。ペニシリン G アシラーゼと呼ばれる酵素は、2 つの中間体である6-アミノペニシラン酸を生成する直接的な加水分解反応を促進します。 (6-APA )およびフェニル酢酸。加水分解に使用される水分子は水素原子とヒドロキシル基に分割され、それぞれ 6-APA とフェニル酢酸に結合します。
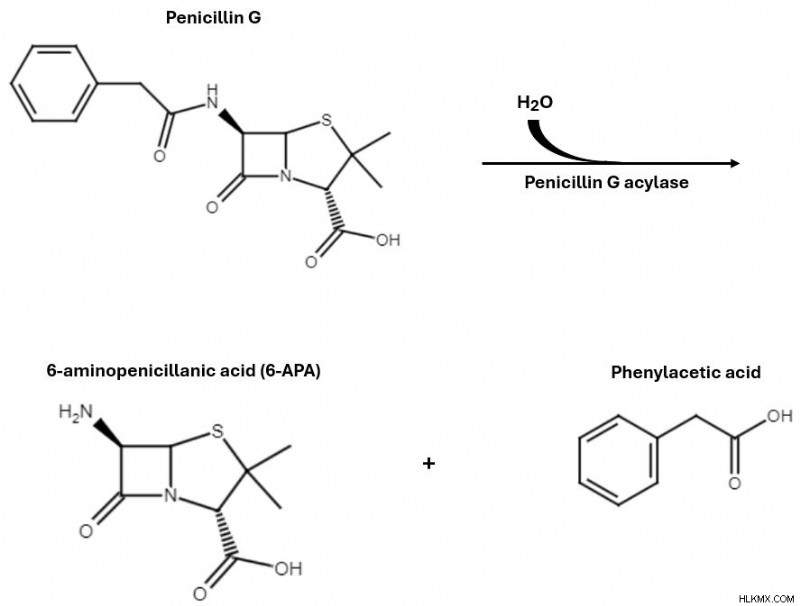 ペニシリン アシラーゼは前駆体としてペニシリン G から始まり、6-APA とフェニル酢酸を生じる加水分解反応を触媒します。
ペニシリン アシラーゼは前駆体としてペニシリン G から始まり、6-APA とフェニル酢酸を生じる加水分解反応を触媒します。 フェニル酢酸は合成の次のステップには関係がないため、この段階では無視します。代わりに、6-APA 中間体の構造が β-ラクタム系抗生物質の構造にどのように似ているかに注目してください。 6-APA を新しい反応物質であるフェノキシ酢酸と組み合わせます。 6-APA は第一級アミンから 1 つの水素原子を失い、フェノキシ酢酸は末端ヒドロキシル基を失います。このステップ中に 1 分子に相当する水分子が除去され、反応によって完全に形成されたペニシリン V 分子が準備されます。
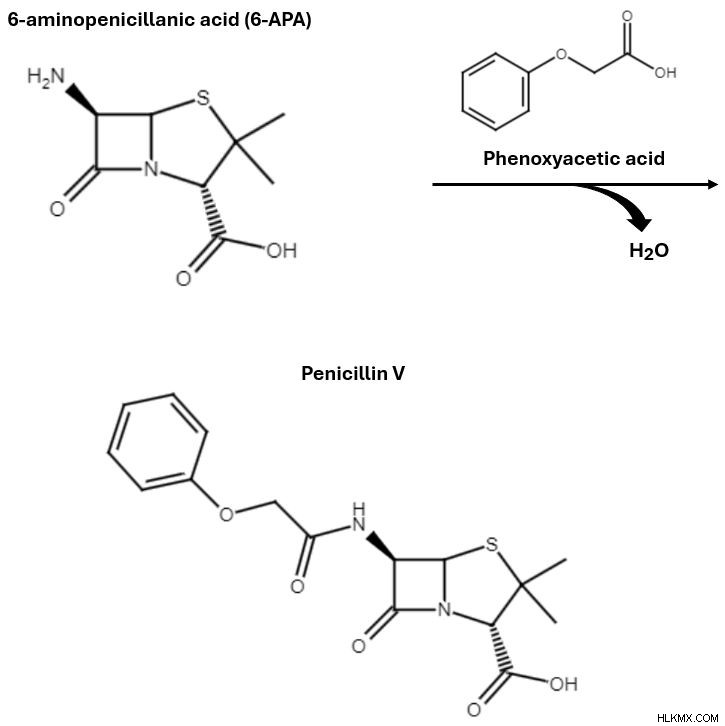 フェノキシ酢酸は 6-APA とアミド結合を形成し、側鎖として特徴的なフェノキシアセチル基を持つペニシリン V を生成します。
フェノキシ酢酸は 6-APA とアミド結合を形成し、側鎖として特徴的なフェノキシアセチル基を持つペニシリン V を生成します。 6-APA を組み込むことには、抗生物質の合成において大きな利点があります。ご覧のとおり、6-APA を目的のペニシリン製品に変換するには、簡単な 1 つのステップのみが必要です。これにより、6-APA は、β-ラクタム系抗生物質を合成する際の有用かつ動的な中間体となります。フェノキシ酢酸を異なる側鎖を持つカルボン酸に置き換えると、異なる R 基を持つペニシリンの形が得られます。このように、自然の生合成と比較して、半合成は、より速く、より簡単で、より多用途なペニシリン生産のための出発点となります。また、アモキシシリンやアンピシリンなど、自然界には決して存在しない抗生物質を製造するための実行可能な選択肢でもあります。
ペニシリンと抗生物質耐性
ペニシリンなどの抗生物質は微妙なバランスで存在します。感染症を治すのに十分な細菌細胞を破壊できますが、すべての細菌細胞を殺すことはできません。抗生物質は、最も攻撃を受けやすい最も弱い細胞を殺します。したがって、生き残った細菌細胞が最も強い細胞になります。これらの強力な細胞は依然として感染を引き起こし、繁殖する可能性があり、この強さは有利な適応であるため、娘細胞にも受け継がれることがよくあります。最終的に、この細菌集団は主に抗生物質による治療に耐えられる細胞で構成されます。
この適応は、医学的なジレンマを引き起こします。抗生物質は重篤な病気を治すことができますが、抗生物質がそれらを殺すことができないほど強力な細菌を発生させる可能性もあります。これらの強力な細菌は耐性があります。 抗生物質に。現代の主要な課題は、抗生物質耐性を最小限に抑えながら効果を最大化する方法で抗生物質を使用する方法です。これが、さまざまな形態のペニシリンを用意することが重要である理由です。利用できる抗生物質の種類が多ければ多いほど、そのうちの少なくとも 1 つが効果を発揮する可能性が高くなります。
幸いなことに、新しい抗生物質の探索は進行中です。そのうちのいくつかは、耐性菌によって引き起こされる感染症を治療するための効果的な選択肢となる可能性があります。新しい抗生物質はどうやって特定するのでしょうか?意図せずに発見されるものもある(フレミングがペニシリンを発見したように)。他の抗生物質研究には、土壌から微生物を分離し、それらの化合物の抗菌活性をスクリーニングすることが含まれます。あるいは、生物医薬品科学者は、新しい抗生物質を作成するために、合成戦略を使用して既知の抗生物質を改変します。これらすべての取り組みが、現在治療不可能な細菌感染症の治療法を提供するという最終目標を推進します。
抗生物質耐性はどのように作用しますか?
抗生物質に耐性を持つためには、細菌細胞が抗生物質の攻撃メカニズムを無効化または無力化する何らかの特性を持っていなければなりません。例として、β-ラクタム系抗生物質耐性の背後にあるメカニズムを調べてみましょう。
B-ラクタマーゼ
ご存知のとおり、β-ラクタム環はペプチドグリカン層の形成を妨げることにより細菌を殺す役割を果たしています。嫌気性細菌はβ- ラクタマーゼを生成します。 、β-ラクタム環のアミド結合を加水分解する酵素。 B-ラクタム環は環に大きな歪みがあり、非常に反応性が高いため、加水分解を受けやすくなります。無傷のβ-ラクタム環がなければ、抗生物質は細菌の細胞壁を標的にすることができません。この場合、抗生物質を投与しても細菌は影響を受けません。
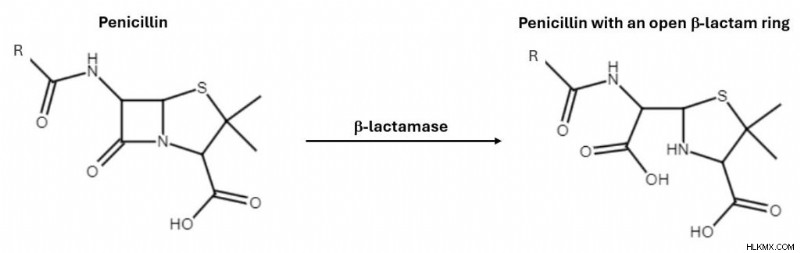
ただし、β-ラクタマーゼ阻害剤は抗生物質と併用して投与することができます。 β-ラクタム系抗生物質とβ-ラクタマーゼ阻害剤を強力に組み合わせると、抗生物質を単独で使用する場合に比べて、耐性菌を倒す可能性が高くなります。 β-ラクタマーゼ阻害剤は、β-ラクタマーゼに不可逆的に結合して不活化することによりアンタゴニストとして機能します。 β-ラクタマーゼが抗生物質のβ-ラクタム環を妨げなくなったので、抗生物質は正常に作用することができます。広く入手可能な β-ラクタマーゼ阻害剤の例には、クラブラン酸やスルバクタムなどがあります。
ペニシリン結合タンパク質
ペニシリン結合タンパク質 (PBP )ペプチドグリカン層の形成を担う反応を触媒します。したがって、PBP を阻害すると細菌の細胞壁が変形または弱くなり、最終的には溶解が起こります。ペプチドグリカンに架橋を形成する酵素、DD トランスペプチダーゼを思い出してください。これは PBP の 1 つです。
PBP は、β-ラクタム環のアミド結合でペニシリンに結合し、β-ラクタマーゼと同様に強制的に環を開くことができます。この結合により PBP が不可逆的に不活性化され、ペニシリンが無効になります。 PBP を発現する細菌の注目すべき例の 1 つは、 メチシリン耐性の黄色ブドウ球菌です。 (MRSA)、メチシリンはペニシリンの一種です。
そのため、抗生物質耐性の解決に努めている科学者の間では、PBP がもう 1 つの研究対象となっています。 PBP を克服すると、ペプチドグリカンの合成が妨げられ、ペニシリンが細菌細胞を攻撃しやすくなります。最近の研究では、PBP の構造、特にその中のアミノ酸を変更することが、ペニシリンと PBP の結合にどのような影響を与えるかを調査しています。 PBP がペニシリンの β-ラクタム環に結合できないように変更された場合、PBP はペニシリンの抗生物質機能を妨げなくなります。
結論
アレクサンダー・フレミングによるペニシリンの予期せぬ発見は、微生物研究の流れを変えました。 β-ラクタム系抗生物質は、β-ラクタム環を使用して DD-トランスペプチダーゼを阻害し、細菌細胞のペプチドグリカンベースの細胞壁を弱めます。ペニシリンには数多くのバリエーションがあり、生合成や半合成など、ペニシリンを生成する方法も数多くあります。
β-ラクタム系抗生物質、その合成、応用、抗生物質耐性の役割についてのダイナミックなディスカッションを掘り下げてみましょう。